

Functional specificity of extracellular nucleases of Shewanella oneidensis MR-1
- PMID: 22492434
- PMCID: PMC3370543
- DOI: 10.1128/AEM.07895-11
Functional specificity of extracellular nucleases of Shewanella oneidensis MR-1
Abstract
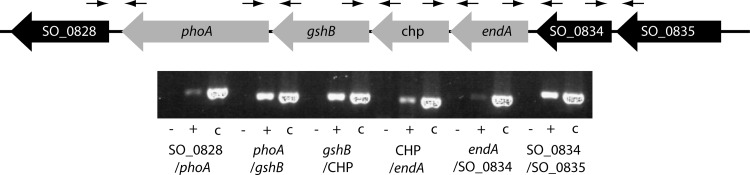
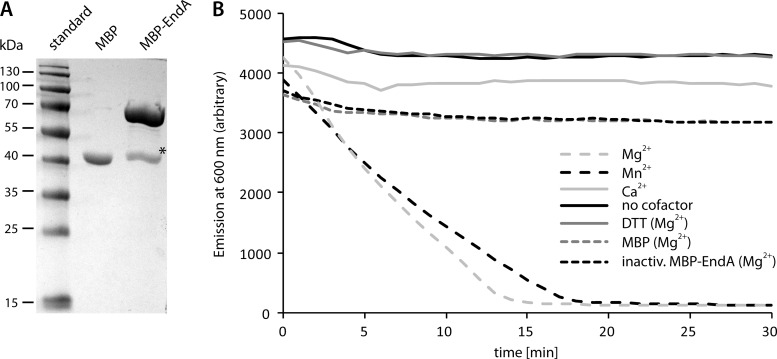
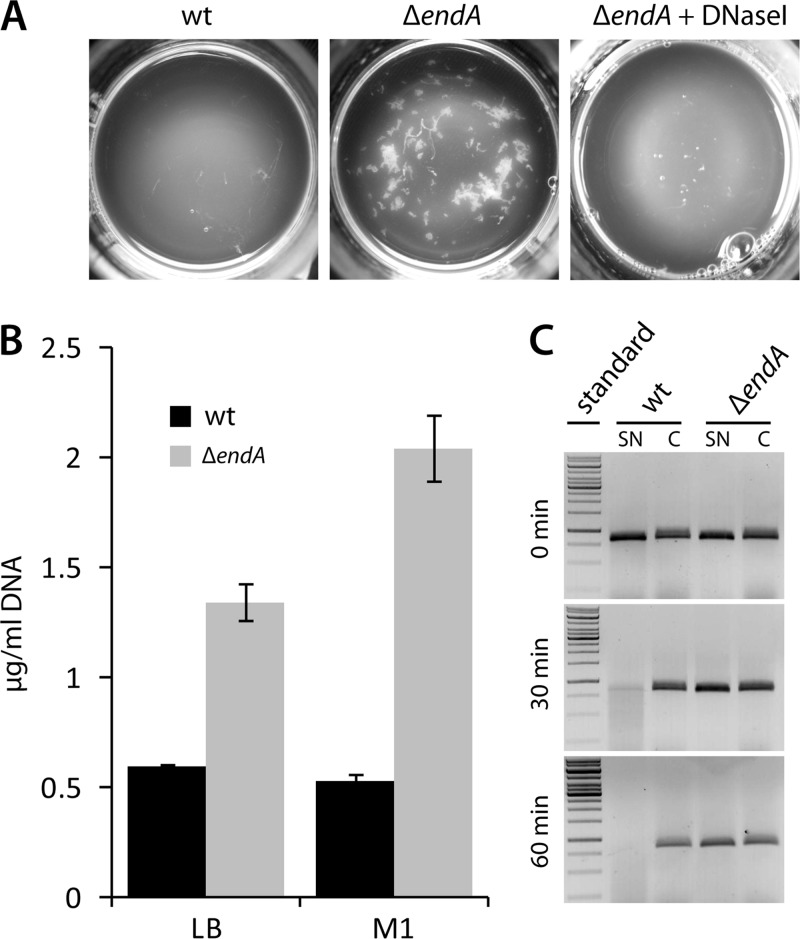
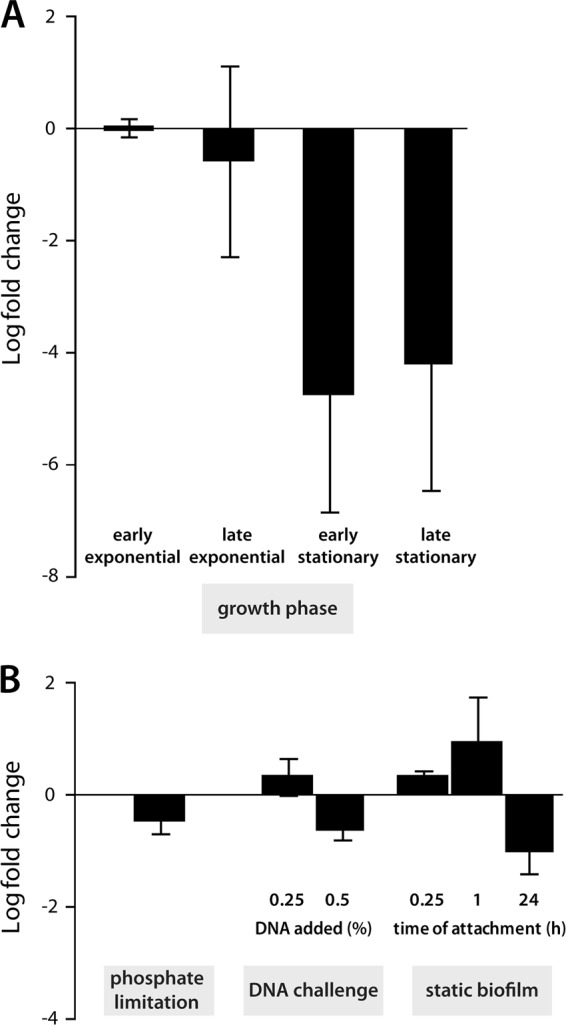
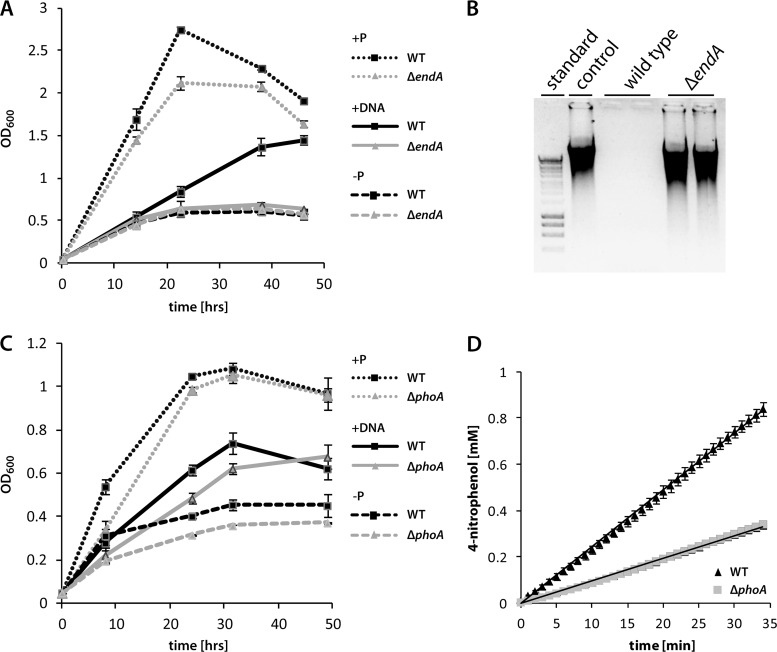
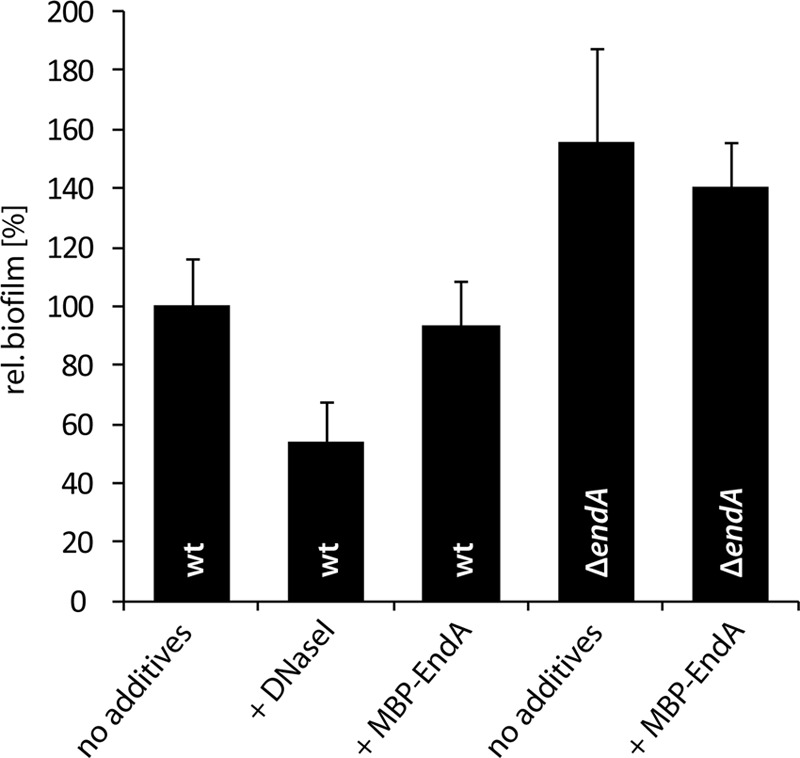
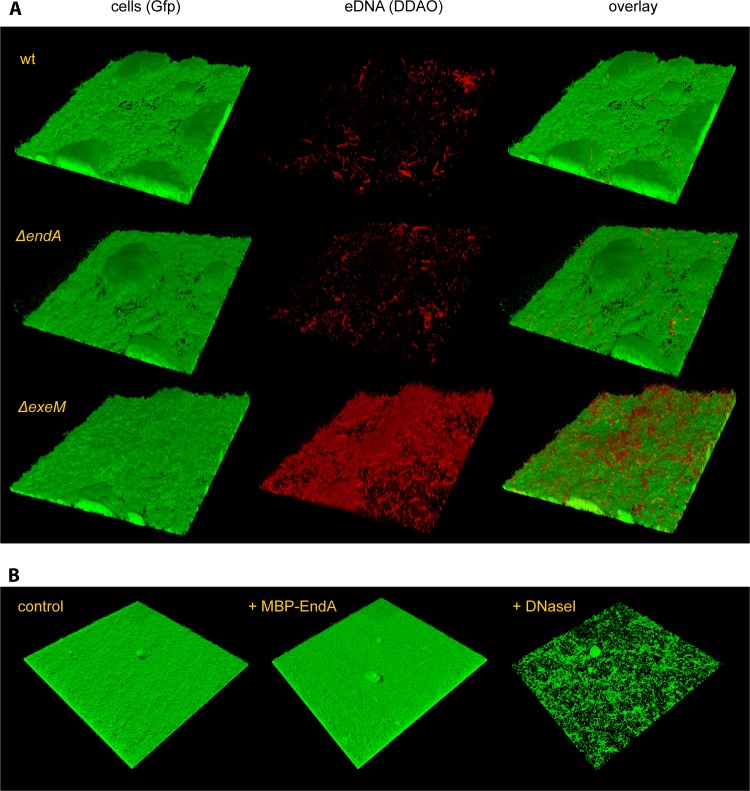
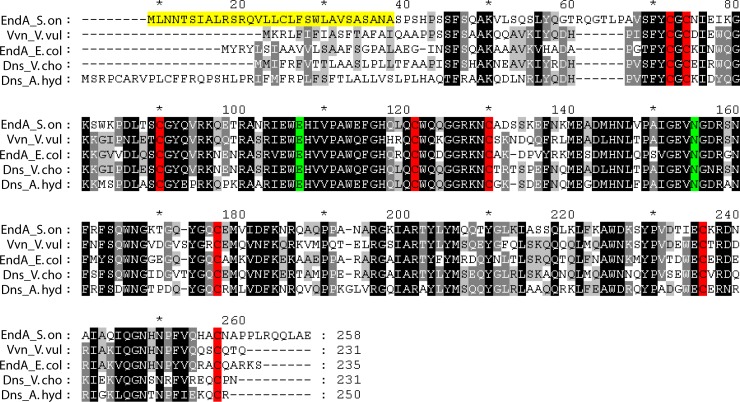
Bacterial species such as Shewanella oneidensis MR-1 require extracellular nucleolytic activity for the utilization of extracellular DNA (eDNA) as a source of nutrients and for the turnover of eDNA as a structural matrix component during biofilm formation. We have previously characterized two extracellular nucleases of S. oneidensis MR-1, ExeM and ExeS. Although both are involved in biofilm formation, they are not specifically required for the utilization of eDNA as a nutrient. Here we identified and characterized EndA, a third extracellular nuclease of Shewanella. The heterologously overproduced and purified protein was highly active and rapidly degraded linear and supercoiled DNAs of various origins. Divalent metal ions (Mg(2+) or Mn(2+)) were required for function. endA is cotranscribed with phoA, an extracellular phosphatase, and is not upregulated upon phosphostarvation. Deletion of endA abolished both extracellular degradation of DNA by S. oneidensis MR-1 and the ability to use eDNA as a sole source of phosphorus. PhoA is not strictly required for the exploitation of eDNA as a nutrient. The activity of EndA prevents the formation of large cell aggregates during planktonic growth. However, in contrast to the findings for ExeM, endA deletion had only minor effects on biofilm formation. The findings strongly suggest that the extracellular nucleases of S. oneidensis exert specific functions required under different conditions.
Figures
References
-
- Aiba H, Adhya S, de Crombrugghe B. 1981. Evidence for two functional gal promoters in intact Escherichia coli cells. J. Biol. Chem. 256:11905–11910 - PubMed
-
- Altermark B, Smalas AO, Willassen NP, Helland R. 2006. The structure of Vibrio cholerae extracellular endonuclease I reveals the presence of a buried chloride ion. Acta Crystallogr. D Biol. Crystallogr. 62:1387–1391 - PubMed
-
- Baldwin DS. 1998. Reactive “organic” phosphorus revisited. Water Res. 32:2265–2270
-
- Bendtsen JD, Nielsen H, von Heijne G, Brunak S. 2004. Improved prediction of signal peptides: SignalP 3.0. J. Mol. Biol. 340:783–795 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
