

Endometrial miR-181a and miR-98 expression is altered during transition from normal into cancerous state and target PGR, PGRMC1, CYP19A1, DDX3X, and TIMP3
- PMID: 22492871
- PMCID: PMC3387391
- DOI: 10.1210/jc.2012-1018
Endometrial miR-181a and miR-98 expression is altered during transition from normal into cancerous state and target PGR, PGRMC1, CYP19A1, DDX3X, and TIMP3
Abstract
Context: Evidence suggests that a number of microRNA (miRNA) are aberrantly expressed in endometrial disorders with potential posttranscriptional regulation of their specific target genes, including ovarian steroid receptors.
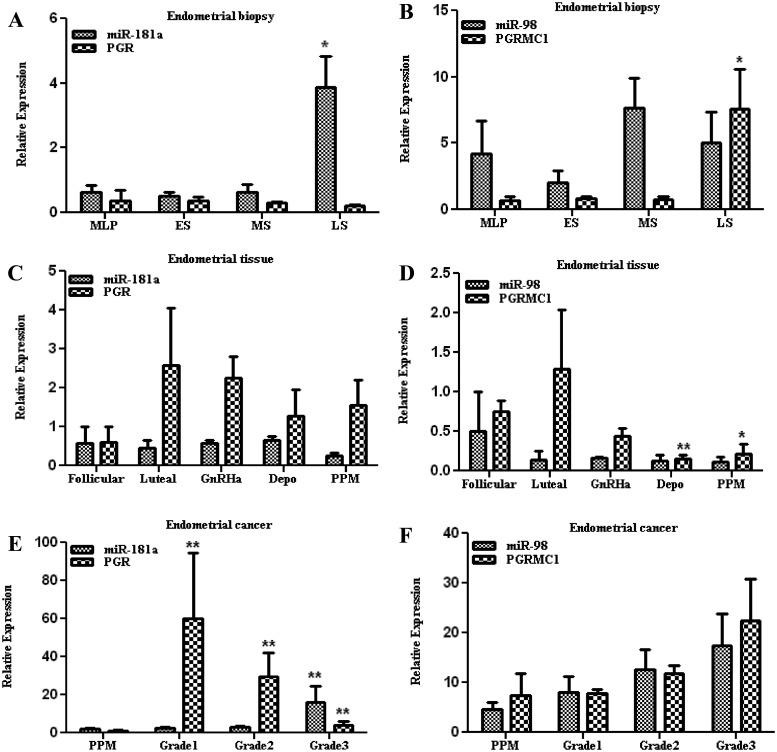
Objectives: Our objective was to assess the endometrial expression of miR-98 and miR-181a and their respective target genes, progesterone (P4) receptor membrane component 1 (PGRMC1) and P4 receptor (PGR).
Design, setting, and patients: We evaluated tissue expression and in vitro regulation at an academic university medical center in endometrial biopsies and endometrial tissues from follicular and luteal phases with and without exposure to hormonal therapies and grade I-III endometrial cancer (n = 52).
Interventions: INTERVENTIONS included endometrial biopsies and in vitro transfection.
Main outcome measures: We evaluated expression and function of miR-98 and miR-181a.
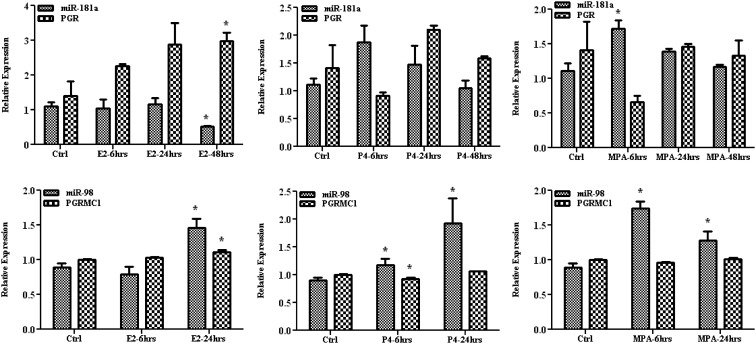
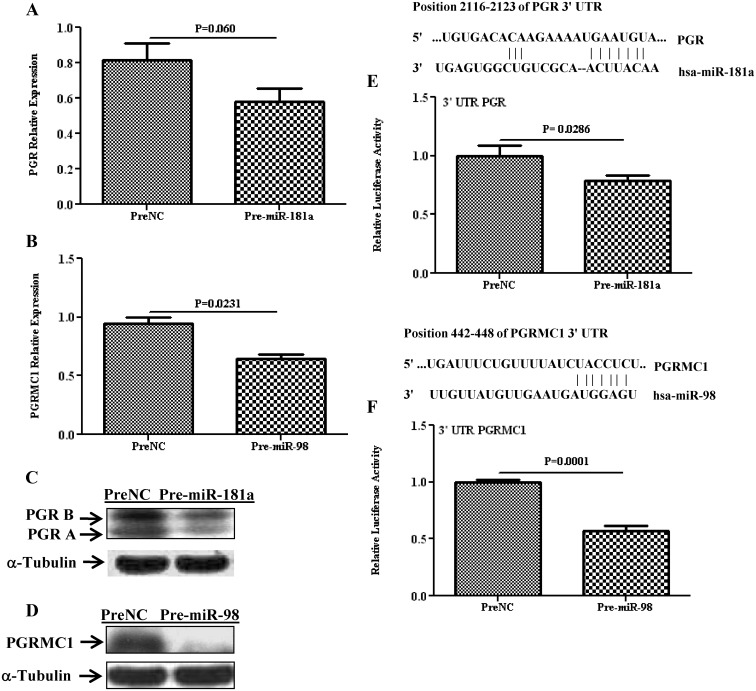
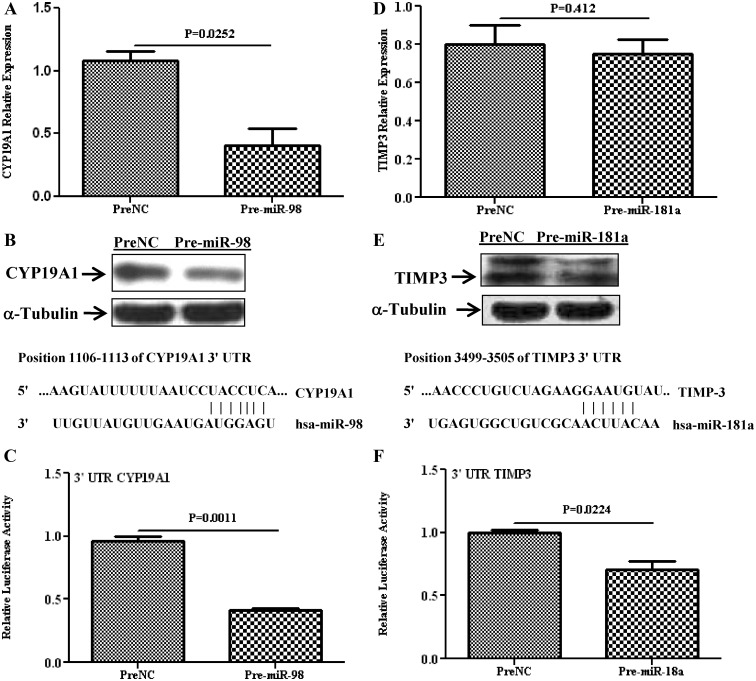
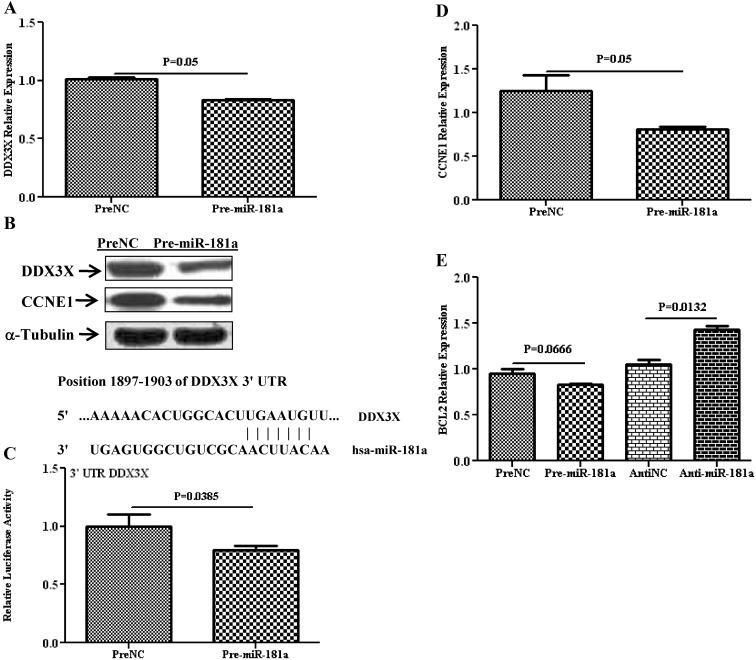
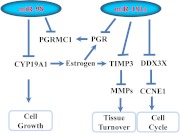
Results: Aberrant expression of miR-98 and miR-181a is associated with endometrial transition from normal into cancerous states, which to some extent is influenced by hormonal milieu, and exhibited an inverse relationship with PGMRC1 and PGR expression, respectively. Treatments of Ishikawa cells with 17β-estradiol, P4, or medroxyprogesterone acetate had limited effects on miR-98, miR-181a, and PGRMC1 expression, whereas 17β-estradiol treatment increased PGR expression. In Ishikawa cells, gain of function of miR-98 repressed PGRMC1 and CYP19A1, and miR-181a repressed PGR, DDX3X, and TIMP3 at mRNA and protein levels through direct interactions with their respective 3'-untranslated regions and CCNE1 through miR-181a-induced DDX3X repression, with miR-98 reducing the rate of cell proliferation as compared with controls.
Conclusion: miR-98 and miR-181a through their regulatory functions on PGRMC1, PGR, CYP19A1, TIMP3, and DDX3X expression may influence a wide range of endometrial cellular activities during normal menstrual cycle and transition into disease states, including endometrial cancer.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
