

ErbB receptor tyrosine kinase/NF-κB signaling controls mammosphere formation in human breast cancer
- PMID: 22492965
- PMCID: PMC3340023
- DOI: 10.1073/pnas.1113271109
ErbB receptor tyrosine kinase/NF-κB signaling controls mammosphere formation in human breast cancer
Abstract
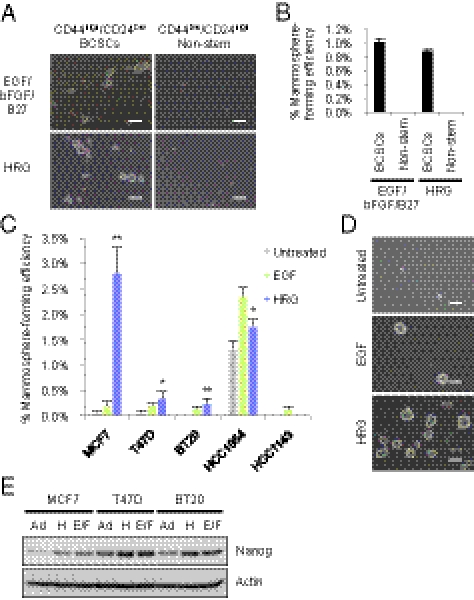
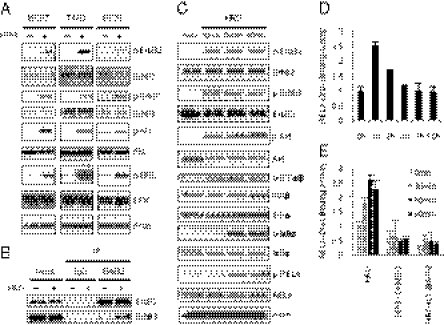
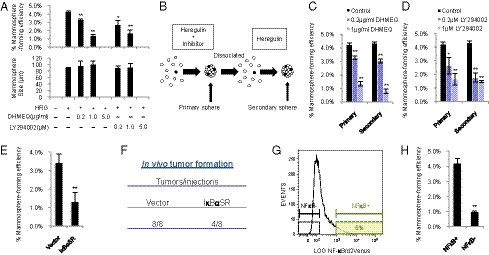
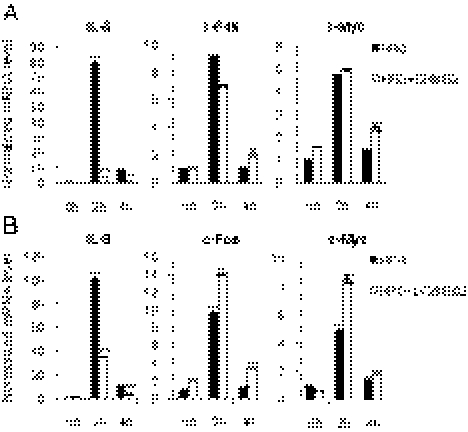
Breast cancer is one of the most common cancers in humans. However, our understanding of the cellular and molecular mechanisms underlying tumorigenesis in breast tissues is limited. Here, we identified a molecular mechanism that controls the ability of breast cancer cells to form multicellular spheroids (mammospheres). We found that heregulin (HRG), a ligand for ErbB3, induced mammosphere formation of a breast cancer stem cell (BCSC)-enriched population as well as in breast cancer cell lines. HRG-induced mammosphere formation was reduced by treatment with inhibitors for phosphatidyl inositol 3-kinase (PI3K) or NF-κB and by expression of IκBα-Super Repressor (IκBαSR), a dominant-negative inhibitor for NF-κB. Moreover, the overexpression of IκBαSR in breast cancer cells inhibited tumorigenesis in NOD/SCID mice. Furthermore, we found that the expression of IL8, a regulator of self-renewal in BCSC-enriched populations, was induced by HRG through the activation of the PI3K/NF-κB pathway. These findings illustrate that HRG/ErbB3 signaling appears to maintain mammosphere formation through a PI3K/NF-κB pathway in human breast cancer.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Reya T, Morrison SJ, Clarke MF, Weissman IL. Stem cells, cancer, and cancer stem cells. Nature. 2001;414:105–111. - PubMed
-
- Gotoh N. Potential signaling pathways activated in cancer stem cells in breast cancer. In: Shostak S, editor. Cancer Stem Cells Theories and Practice. Vienna, Austria: InTech; 2011. p. 6.
-
- Bonnet D, Dick JE. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat Med. 1997;3:730–737. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
