

Characterization of cleavage events in the multifunctional cilium adhesin Mhp684 (P146) reveals a mechanism by which Mycoplasma hyopneumoniae regulates surface topography
- PMID: 22493032
- PMCID: PMC3322551
- DOI: 10.1128/mBio.00282-11
Characterization of cleavage events in the multifunctional cilium adhesin Mhp684 (P146) reveals a mechanism by which Mycoplasma hyopneumoniae regulates surface topography
Abstract
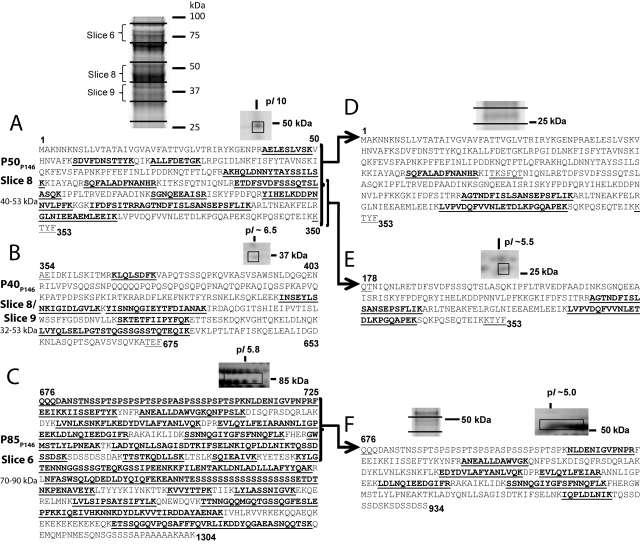
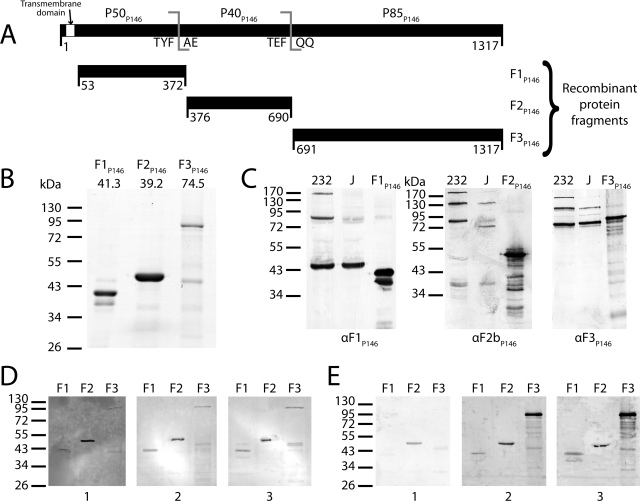
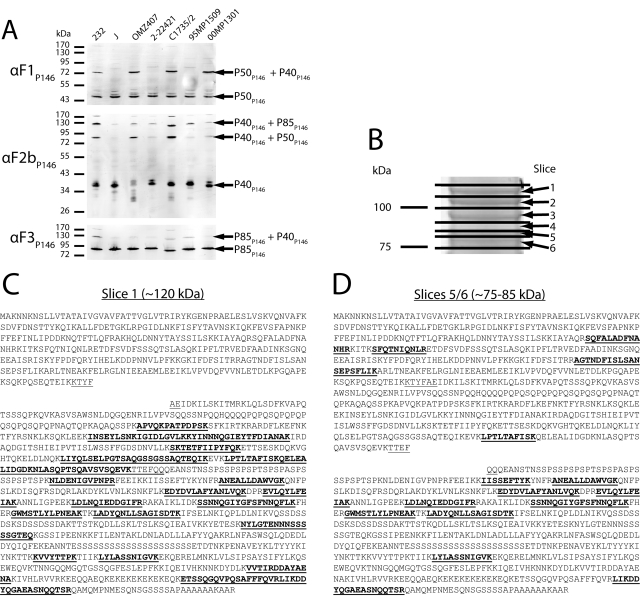
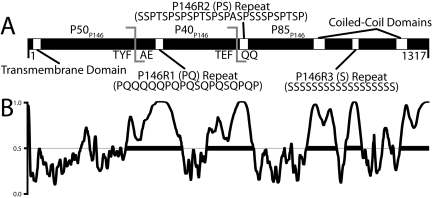
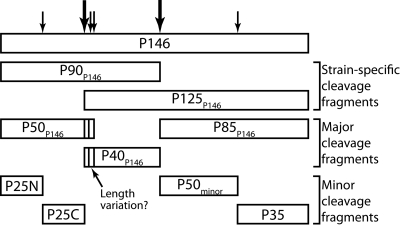
Mycoplasma hyopneumoniae causes enormous economic losses to swine production worldwide by colonizing the ciliated epithelium in the porcine respiratory tract, resulting in widespread damage to the mucociliary escalator, prolonged inflammation, reduced weight gain, and secondary infections. Protein Mhp684 (P146) comprises 1,317 amino acids, and while the N-terminal 400 residues display significant sequence identity to the archetype cilium adhesin P97, the remainder of the molecule is novel and displays unusual motifs. Proteome analysis shows that P146 preprotein is endogenously cleaved into three major fragments identified here as P50(P146), P40(P146), and P85(P146) that reside on the cell surface. Liquid chromatography with tandem mass spectrometry (LC-MS/MS) identified a semitryptic peptide that delineated a major cleavage site in Mhp684. Cleavage occurred at the phenylalanine residue within sequence (672)ATEF↓QQ(677), consistent with a cleavage motif resembling S/T-X-F↓X-D/E recently identified in Mhp683 and other P97/P102 family members. Biotinylated surface proteins recovered by avidin chromatography and separated by two-dimensional gel electrophoresis (2-D GE) showed that more-extensive endoproteolytic cleavage of P146 occurs. Recombinant fragments F1(P146)-F3(P146) that mimic P50(P146), P40(P146), and P85(P146) were constructed and shown to bind porcine epithelial cilia and biotinylated heparin with physiologically relevant affinity. Recombinant versions of F3(P146) generated from M. hyopneumoniae strain J and 232 sequences strongly bind porcine plasminogen, and the removal of their respective C-terminal lysine and arginine residues significantly reduces this interaction. These data reveal that P146 is an extensively processed, multifunctional adhesin of M. hyopneumoniae. Extensive cleavage coupled with variable cleavage efficiency provides a mechanism by which M. hyopneumoniae regulates protein topography.
Importance: Vaccines used to control Mycoplasma hyopneumoniae infection provide only partial protection. Proteins of the P97/P102 families are highly expressed, functionally redundant molecules that are substrates of endoproteases that generate multifunctional adhesin fragments on the cell surface. We show that P146 displays a chimeric structure consisting of an N terminus, which shares sequence identity with P97, and novel central and C-terminal regions. P146 is endoproteolytically processed at multiple sites, generating at least nine fragments on the surface of M. hyopneumoniae. Dominant cleavage events occurred at S/T-X-F↓X-D/E-like sites generating P50(P146), P40(P146), and P85(P146). Recombinant proteins designed to mimic the major cleavage fragments bind porcine cilia, heparin, and plasminogen. P146 undergoes endoproteolytic processing events at multiple sites and with differential processing efficiency, generating combinatorial diversity on the surface of M. hyopneumoniae.
Figures
References
-
- Browning GF, Marenda MS, Noormohammadi AH, Markham PF. 2011. The central role of lipoproteins in the pathogenesis of mycoplasmoses. Vet. Microbiol. 153:44–50 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
