

PERK utilizes intrinsic lipid kinase activity to generate phosphatidic acid, mediate Akt activation, and promote adipocyte differentiation
- PMID: 22493067
- PMCID: PMC3372262
- DOI: 10.1128/MCB.00063-12
PERK utilizes intrinsic lipid kinase activity to generate phosphatidic acid, mediate Akt activation, and promote adipocyte differentiation
Abstract
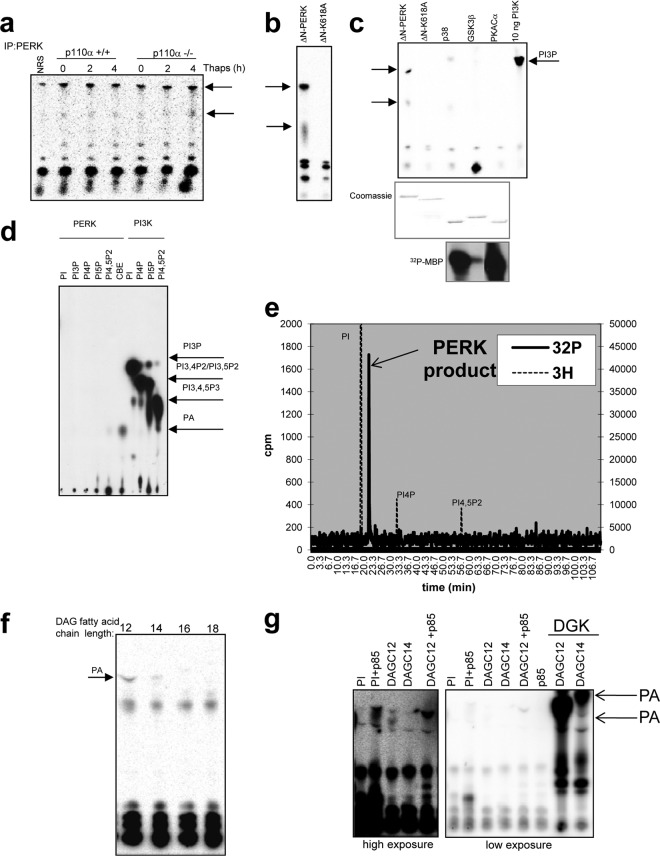
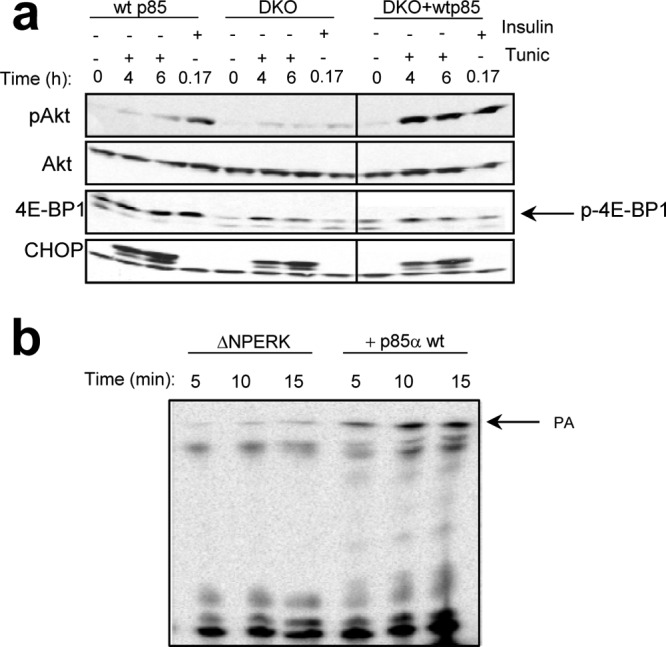
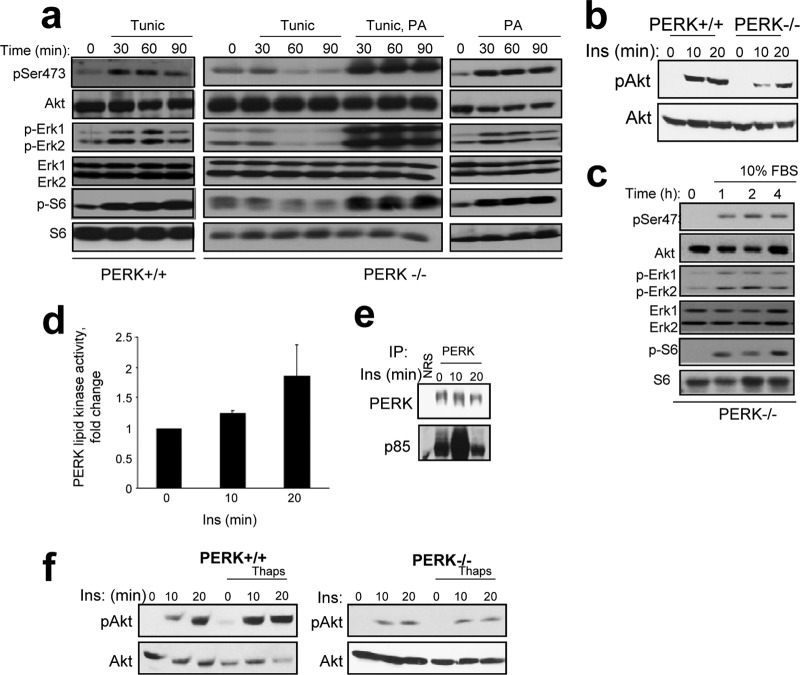
The endoplasmic reticulum (ER) resident PKR-like kinase (PERK) is necessary for Akt activation in response to ER stress. We demonstrate that PERK harbors intrinsic lipid kinase, favoring diacylglycerol (DAG) as a substrate and generating phosphatidic acid (PA). This activity of PERK correlates with activation of mTOR and phosphorylation of Akt on Ser473. PERK lipid kinase activity is regulated in a phosphatidylinositol 3-kinase (PI3K) p85α-dependent manner. Moreover, PERK activity is essential during adipocyte differentiation. Because PA and Akt regulate many cellular functions, including cellular survival, proliferation, migratory responses, and metabolic adaptation, our findings suggest that PERK has a more extensive role in insulin signaling, insulin resistance, obesity, and tumorigenesis than previously thought.
Figures




References
-
- Antonsson B. 1997. Phosphatidylinositol synthase from mammalian tissues. Biochim. Biophys. Acta 1348: 179– 186 - PubMed
-
- Avila-Flores A, Santos T, Rincon E, Merida I. 2005. Modulation of the mammalian target of rapamycin pathway by diacylglycerol kinase-produced phosphatidic acid. J. Biol. Chem. 280: 10091– 10099 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous