

Phosphatidylinositol 3-kinase-binding protein, PI3KAP/XB130, is required for cAMP-induced amplification of IGF mitogenic activity in FRTL-5 thyroid cells
- PMID: 22496359
- PMCID: PMC5416991
- DOI: 10.1210/me.2011-1349
Phosphatidylinositol 3-kinase-binding protein, PI3KAP/XB130, is required for cAMP-induced amplification of IGF mitogenic activity in FRTL-5 thyroid cells
Abstract
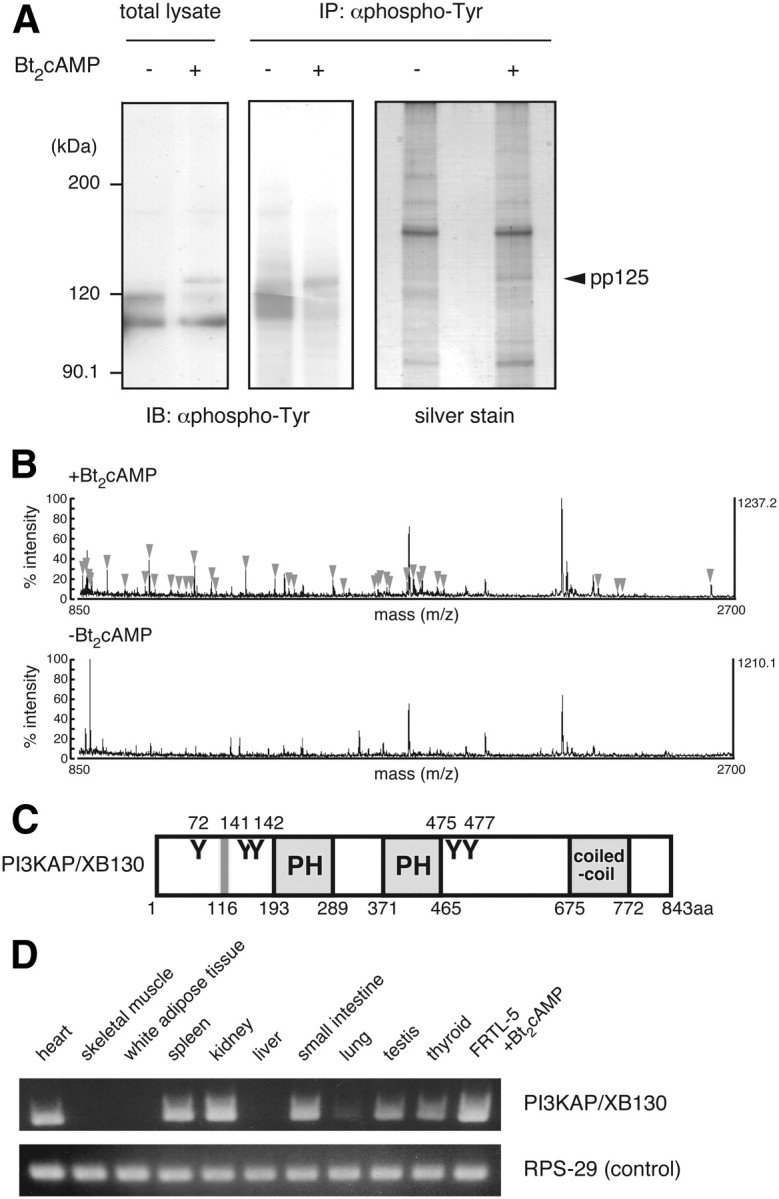
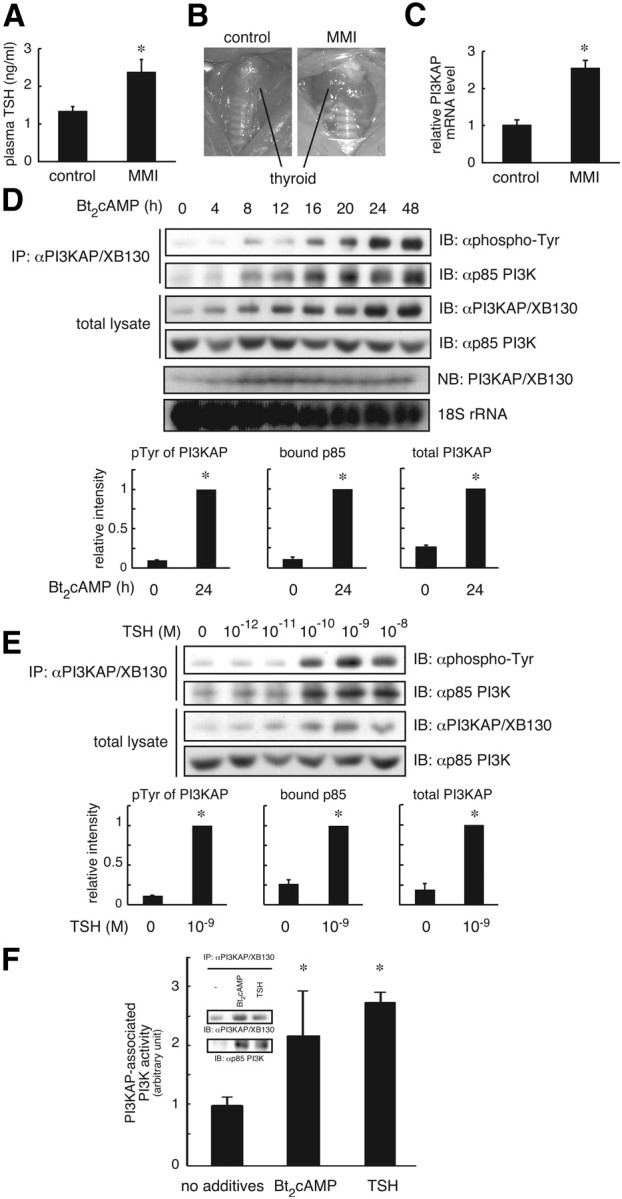
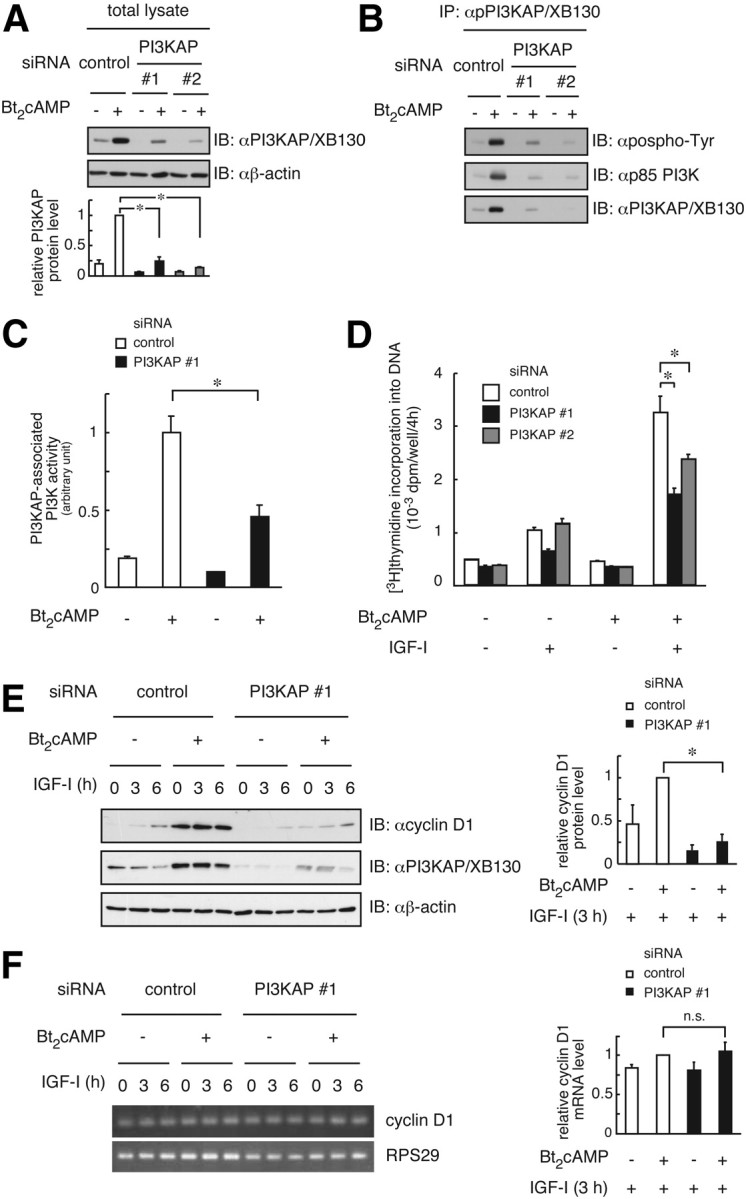
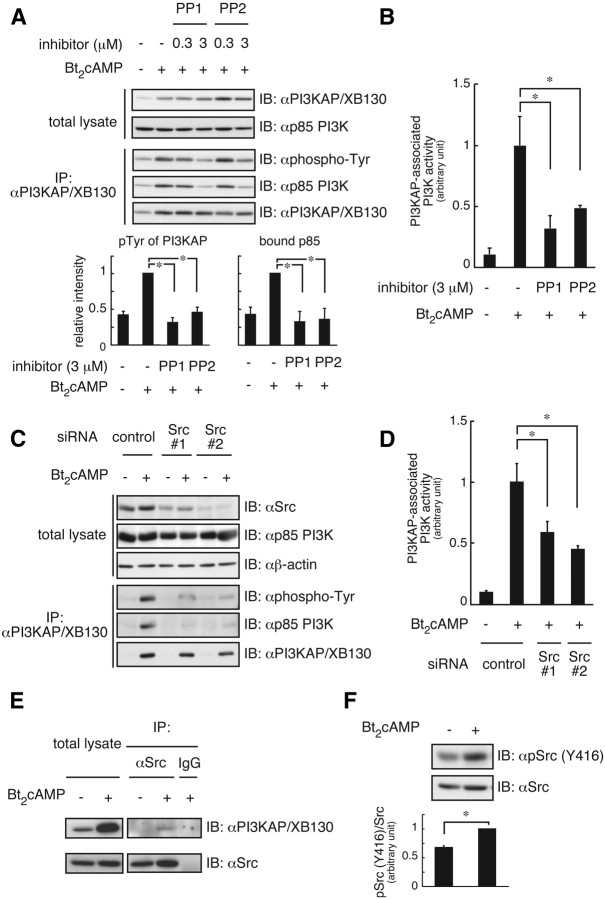
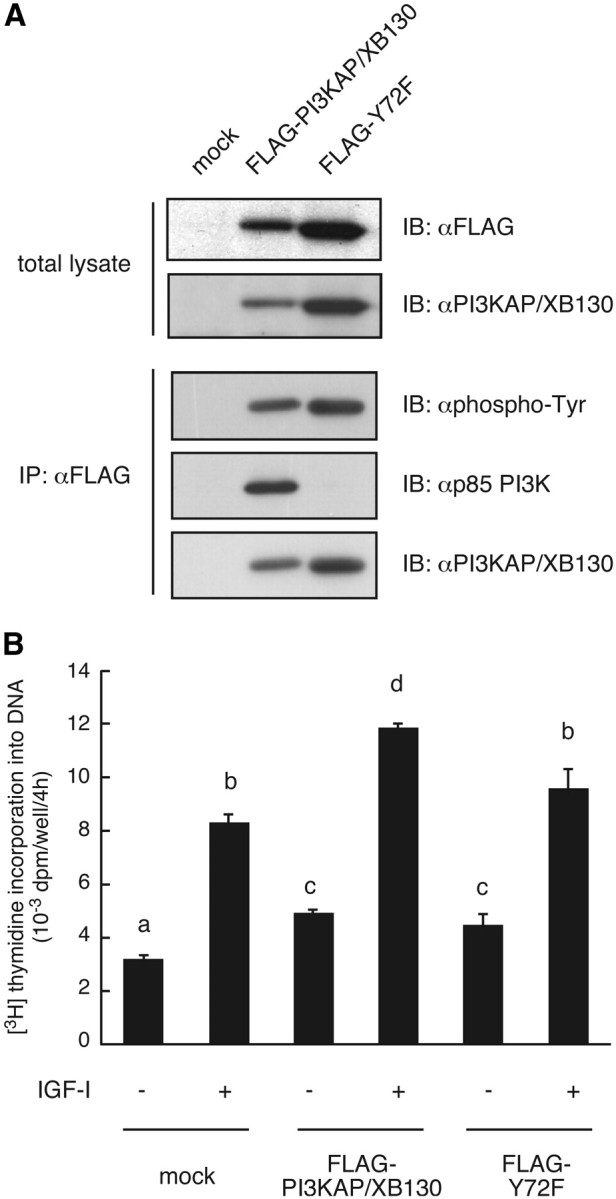
We previously demonstrated that long-term pretreatment of rat FRTL-5 thyroid cells with TSH or cAMP-generating reagents potentiated IGF-I-dependent DNA synthesis. Under these conditions, cAMP treatment increased tyrosine phosphorylation of a 125-kDa protein (p125) and its association with a p85 regulatory subunit of phosphatidylinositol 3-kinase (p85 PI3K), which were suggested to mediate potentiation of DNA synthesis. This study was undertaken to identify p125 and to elucidate its roles in potentiation of DNA synthesis induced by IGF-I. Matrix-assisted laser desorption ionization-time of flight mass spectrometry (MALDI-TOF MS) analysis revealed p125 to be a rat ortholog of human XB130, which we named PI3K-associated protein (PI3KAP). cAMP treatment elevated PI3KAP/XB130 mRNA and protein levels as well as tyrosine phosphorylation and interaction with p85 PI3K leading to increased PI3K activities associated with PI3KAP/XB130, supporting the role of PI3KAP/XB130 in DNA synthesis potentiation. Importantly, PI3KAP/XB130 knockdown attenuated cAMP-dependent potentiation of IGF-I-induced DNA synthesis. Furthermore, c-Src was associated with PI3KAP/XB130 and was activated in response to cAMP. Addition of Src family kinase inhibitors, PP1 or PP2, during cAMP treatment abolished tyrosine phosphorylation of PI3KAP/XB130 and its interaction with p85 PI3K. Finally, introduction of PI3KAP/XB130 into NIH3T3 fibroblasts lacking endogenous PI3KAP/XB130 enhanced IGF-I-induced DNA synthesis; however, a mutant Y72F incapable of binding to p85 PI3K did not show this response. Together, these data indicate that cAMP-dependent induction of PI3KAP/XB130, which is associated with PI3K, is required for enhancement of IGF mitogenic activities.
Figures
Similar articles
-
Tyrosine kinase and phosphatidylinositol 3-kinase activation are required for cyclic adenosine 3',5'-monophosphate-dependent potentiation of deoxyribonucleic acid synthesis induced by insulin-like growth factor-I in FRTL-5 cells.Endocrinology. 2000 Jul;141(7):2429-38. doi: 10.1210/endo.141.7.7539. Endocrinology. 2000. PMID: 10875243
-
Signalling pathways of insulin-like growth factor-I that are augmented by cAMP in FRTL-5 cells.Biochem J. 2000 Jun 1;348 Pt 2(Pt 2):409-16. Biochem J. 2000. PMID: 10816436 Free PMC article.
-
Distinct modes of activation of phosphatidylinositol 3-kinase in response to cyclic adenosine 3', 5'-monophosphate or insulin-like growth factor I play different roles in regulation of cyclin D1 and p27Kip1 in FRTL-5 cells.Endocrinology. 2008 Jul;149(7):3729-42. doi: 10.1210/en.2007-1443. Epub 2008 Apr 10. Endocrinology. 2008. PMID: 18403485
-
Regulation of thyroid cell proliferation by TSH and other factors: a critical evaluation of in vitro models.Endocr Rev. 2001 Oct;22(5):631-56. doi: 10.1210/edrv.22.5.0444. Endocr Rev. 2001. PMID: 11588145 Review.
-
XB130-A Novel Adaptor Protein: Gene, Function, and Roles in Tumorigenesis.Scientifica (Cairo). 2014;2014:903014. doi: 10.1155/2014/903014. Epub 2014 Jun 5. Scientifica (Cairo). 2014. PMID: 24995146 Free PMC article. Review.
Cited by
-
Thyroidal Transcriptomic Profiles of Pathoadaptive Responses to Congenital Hypothyroidism in XB130 Knockout Mice.Cells. 2022 Mar 12;11(6):975. doi: 10.3390/cells11060975. Cells. 2022. PMID: 35326426 Free PMC article.
-
XB130 mediates cancer cell proliferation and survival through multiple signaling events downstream of Akt.PLoS One. 2012;7(8):e43646. doi: 10.1371/journal.pone.0043646. Epub 2012 Aug 23. PLoS One. 2012. PMID: 22928011 Free PMC article.
-
Regulation and functions of MicroRNA-149 in human cancers.Cell Prolif. 2018 Oct;51(5):e12465. doi: 10.1111/cpr.12465. Epub 2018 Jul 12. Cell Prolif. 2018. PMID: 29999552 Free PMC article. Review.
-
XB130 deficiency affects tracheal epithelial differentiation during airway repair.PLoS One. 2014 Oct 1;9(10):e108952. doi: 10.1371/journal.pone.0108952. eCollection 2014. PLoS One. 2014. PMID: 25272040 Free PMC article.
-
Adaptor protein XB130 regulates the aggressiveness of cholangiocarcinoma.PLoS One. 2021 Nov 15;16(11):e0259075. doi: 10.1371/journal.pone.0259075. eCollection 2021. PLoS One. 2021. PMID: 34780466 Free PMC article.
References
-
- Powell-Braxton L , Hollingshead P , Warburton C , Dowd M , Pitts-Meek S , Dalton D , Gillett N , Stewart TA. 1993. IGF-I is required for normal embryonic growth in mice. Genes Dev 7:2609–2617 - PubMed
-
- Baker J , Liu JP , Robertson EJ , Efstratiadis A. 1993. Role of insulin-like growth factors in embryonic and postnatal growth. Cell 75:73–82 - PubMed
-
- Liu JP , Baker J , Perkins AS , Robertson EJ , Efstratiadis A. 1993. Mice carrying null mutations of the genes encoding insulin-like growth factor I (Igf-1) and type 1 IGF receptor (Igf1r). Cell 75:59–72 - PubMed
-
- Jones JI , Clemmons DR. 1995. Insulin-like growth factors and their binding proteins: biological actions. Endocr Rev 16:3–34 - PubMed
-
- Qureshi FG , Tchorzewski MT , Duncan MD , Harmon JW. 1997. EGF and IGF-I synergistically stimulate proliferation of human esophageal epithelial cells. J Surg Res 69:354–358 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Miscellaneous
