

Dopamine modulation of Ih improves temporal fidelity of spike propagation in an unmyelinated axon
- PMID: 22496556
- PMCID: PMC3347488
- DOI: 10.1523/JNEUROSCI.6320-11.2012
Dopamine modulation of Ih improves temporal fidelity of spike propagation in an unmyelinated axon
Abstract
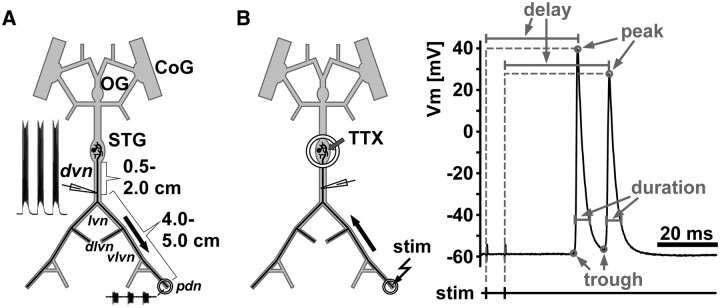
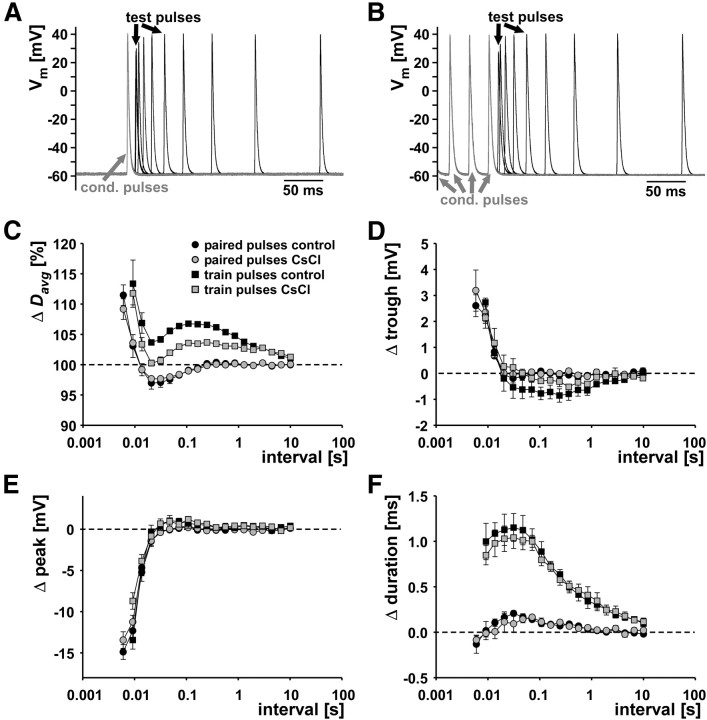
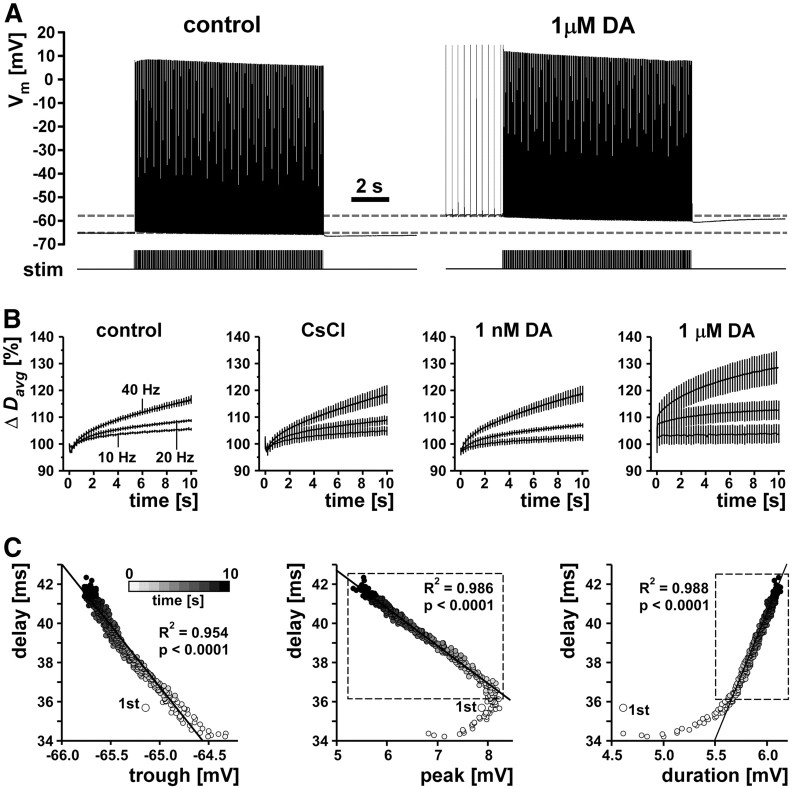
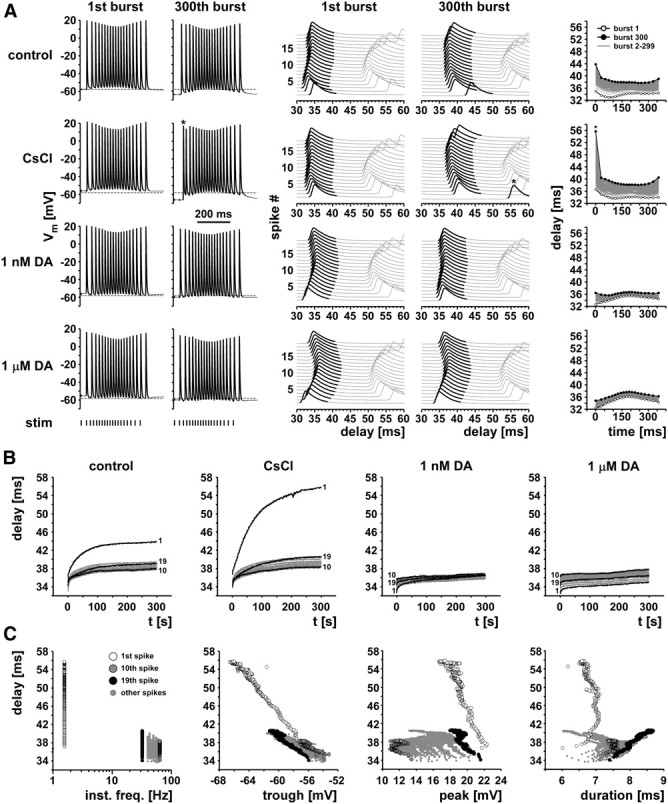
We studied how conduction delays of action potentials in an unmyelinated axon depended on the history of activity and how this dependence was changed by the neuromodulator dopamine (DA). The pyloric dilator axons of the stomatogastric nervous system in the lobster, Homarus americanus, exhibited substantial activity-dependent hyperpolarization and changes in spike shape during repetitive activation. The conduction delays varied by several milliseconds per centimeter, and, during activation with realistic burst patterns or Poisson-like patterns, changes in delay occurred over multiple timescales. The mean delay increased, whereas the resting membrane potential hyperpolarized with a time constant of several minutes. Concomitantly with the mean delay, the variability of delay also increased. The variability of delay was not a linear or monotonic function of instantaneous spike frequency or spike shape parameters, and the relationship between these parameters changed with the increase in mean delay. Hyperpolarization was counteracted by a hyperpolarization-activated inward current (I(h)), and the magnitude of I(h) critically determined the temporal fidelity of spike propagation. Pharmacological block of I(h) increased the change in delay and the variability of delay, and increasing I(h) by application of DA diminished both. Consequently, the temporal fidelity of pattern propagation was substantially improved in DA. Standard measurements of changes in excitability or delay with paired stimuli or tonic stimulation failed to capture the dynamics of spike conduction. These results indicate that spike conduction can be extremely sensitive to the history of axonal activity and to the presence of neuromodulators, with potentially important consequences for temporal coding.
Figures

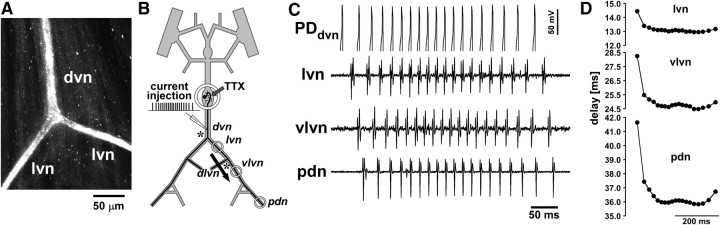
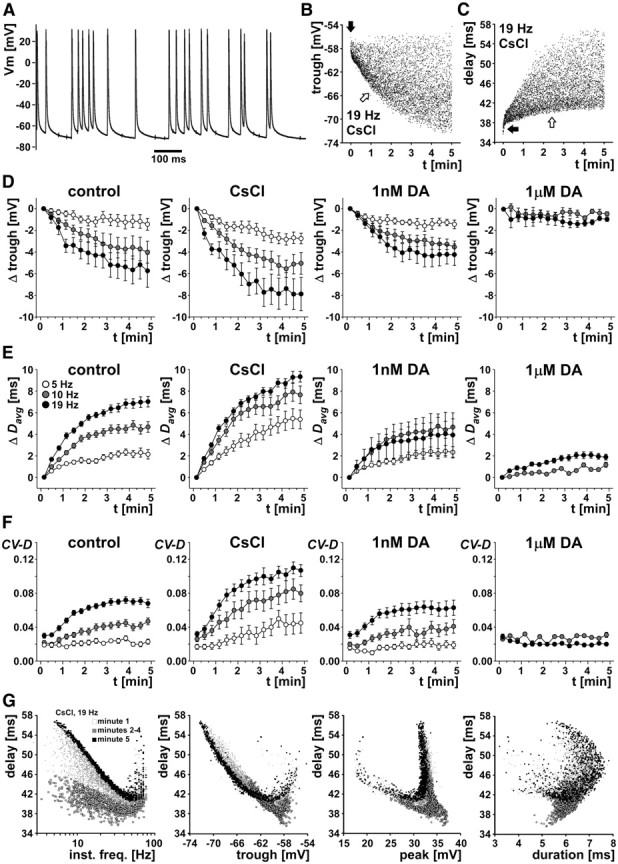
References
-
- Avoli M, Methot M, Kawasaki H. GABA-dependent generation of ectopic action potentials in the rat hippocampus. Eur J Neurosci. 1998;10:2714–2722. - PubMed
-
- Baginskas A, Palani D, Chiu K, Raastad M. The H-current secures action potential transmission at high frequencies in rat cerebellar parallel fibers. Eur J Neurosci. 2009;29:87–96. - PubMed
-
- Baker MD. Axonal flip-flops and oscillators. Trends Neurosci. 2000;23:514–519. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources