

Energetic selection of topology in ferredoxins
- PMID: 22496635
- PMCID: PMC3320576
- DOI: 10.1371/journal.pcbi.1002463
Energetic selection of topology in ferredoxins
Abstract
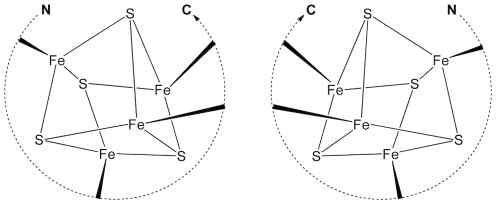
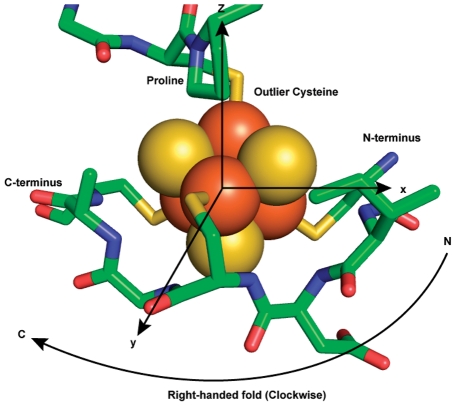
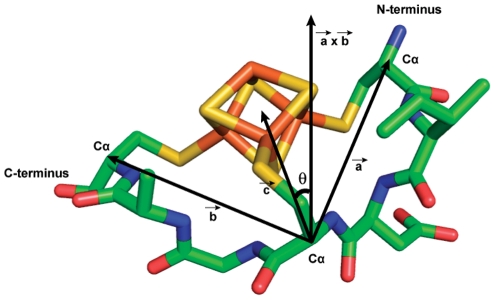
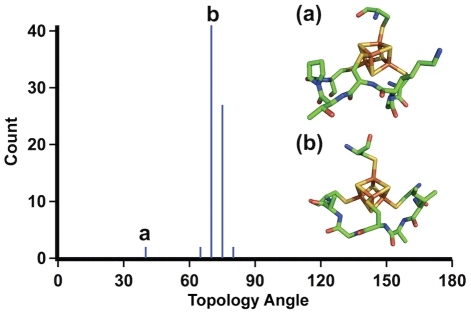
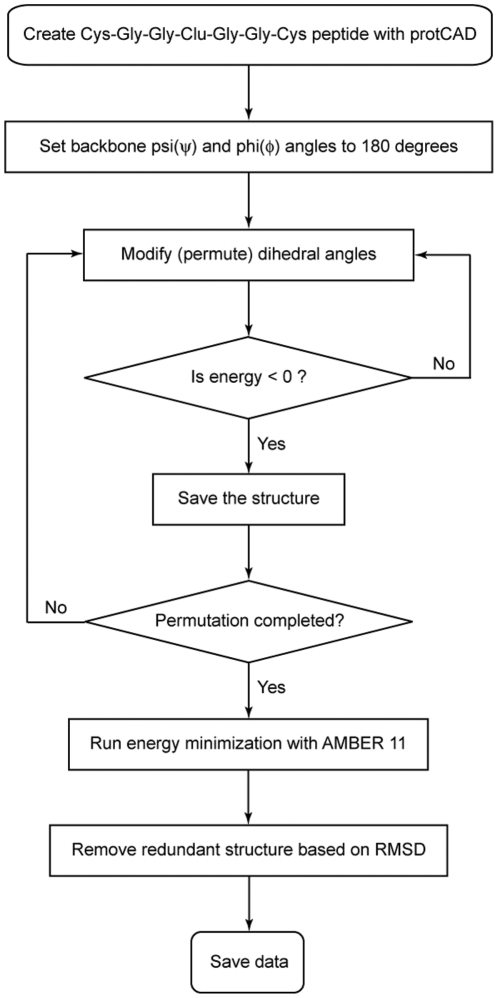
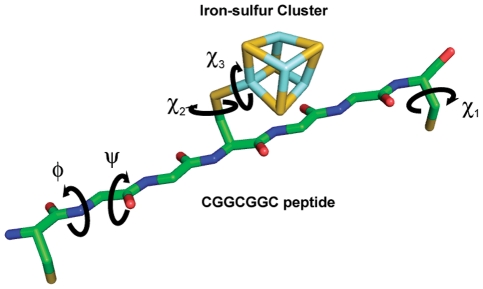
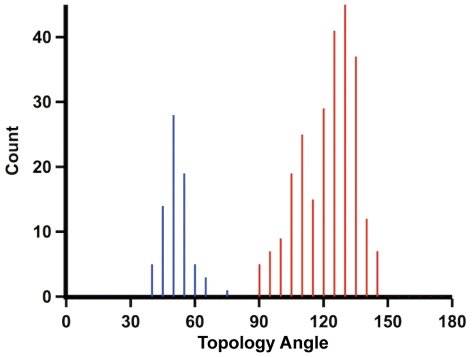
Models of early protein evolution posit the existence of short peptides that bound metals and ions and served as transporters, membranes or catalysts. The Cys-X-X-Cys-X-X-Cys heptapeptide located within bacterial ferredoxins, enclosing an Fe₄S₄ metal center, is an attractive candidate for such an early peptide. Ferredoxins are ancient proteins and the simple α+β fold is found alone or as a domain in larger proteins throughout all three kingdoms of life. Previous analyses of the heptapeptide conformation in experimentally determined ferredoxin structures revealed a pervasive right-handed topology, despite the fact that the Fe₄S₄ cluster is achiral. Conformational enumeration of a model CGGCGGC heptapeptide bound to a cubane iron-sulfur cluster indicates both left-handed and right-handed folds could exist and have comparable stabilities. However, only the natural ferredoxin topology provides a significant network of backbone-to-cluster hydrogen bonds that would stabilize the metal-peptide complex. The optimal peptide configuration (alternating α(L),α(R)) is that of an α-sheet, providing an additional mechanism where oligomerization could stabilize the peptide and facilitate iron-sulfur cluster binding.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Caetano-Anolles G, Caetano-Anolles D, Kim KM, Mittenthal JE. Proteome Evolution and the Metabolic Origins of Translation and Cellular Life. J Mol Evol. 2011;72:14–33. - PubMed
-
- Falkowski PG, Fenchel T, Delong EF. The microbial engines that drive Earth's biogeochemical cycles. Science. 2008;320:1034–1039. - PubMed
-
- Eck RV, Dayhoff MO. Evolution of the structure of ferredoxin based on living relics of primitive amino acid sequences. Science. 1966;152:363–366. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
