

Small-molecule inhibitors of dengue-virus entry
- PMID: 22496653
- PMCID: PMC3320583
- DOI: 10.1371/journal.ppat.1002627
Small-molecule inhibitors of dengue-virus entry
Erratum in
-
Correction: Small-Molecule Inhibitors of Dengue-Virus Entry.PLoS Pathog. 2019 Jan 31;15(1):e1007553. doi: 10.1371/journal.ppat.1007553. eCollection 2019 Jan. PLoS Pathog. 2019. PMID: 30703168 Free PMC article.
Abstract
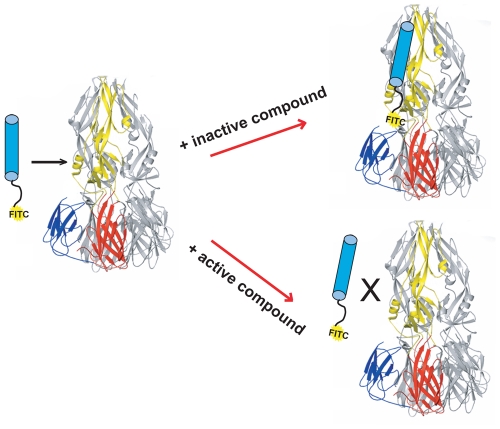
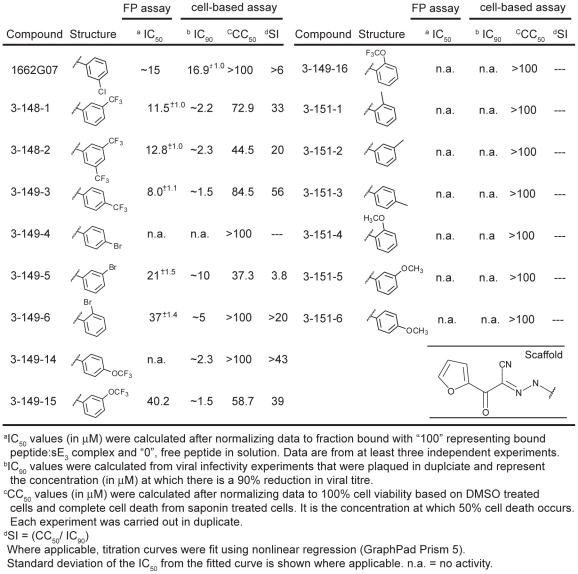
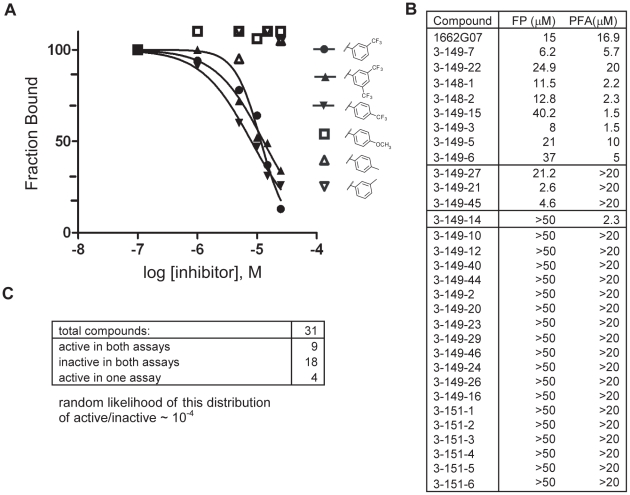
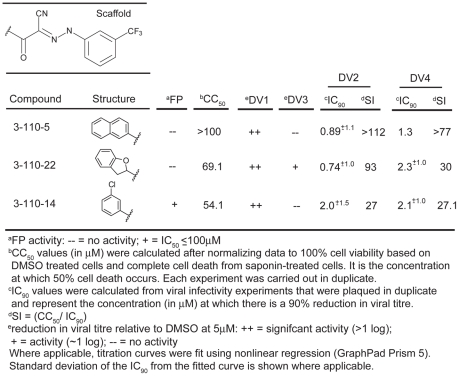
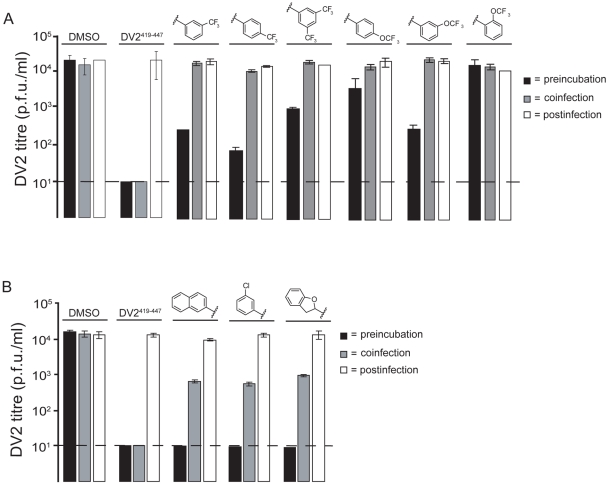
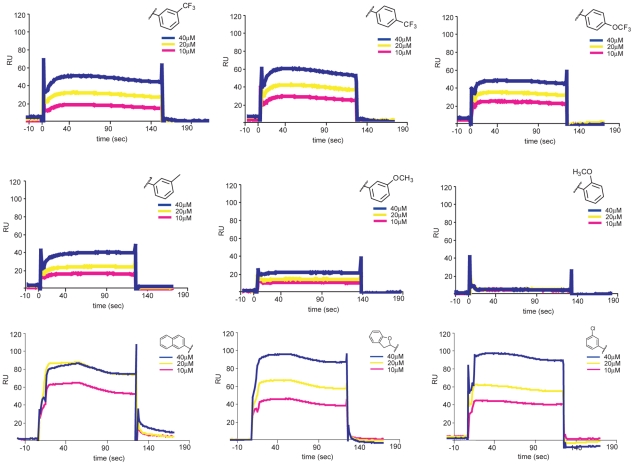
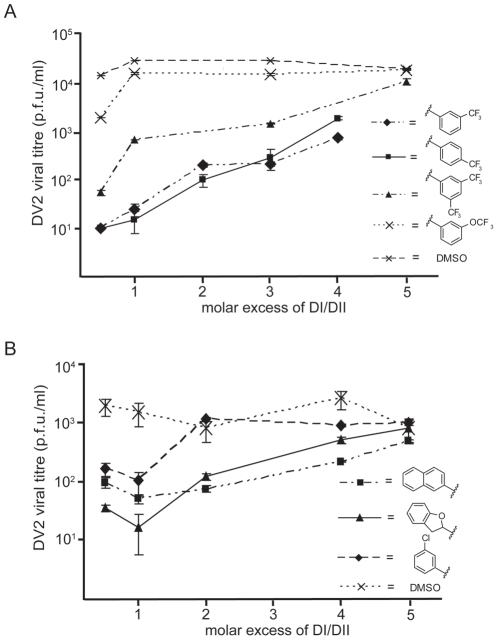
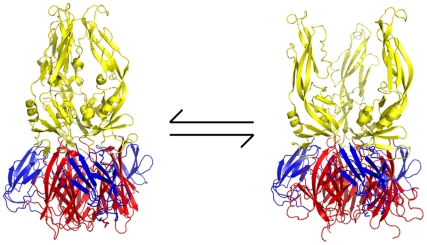
Flavivirus envelope protein (E) mediates membrane fusion and viral entry from endosomes. A low-pH induced, dimer-to-trimer rearrangement and reconfiguration of the membrane-proximal "stem" of the E ectodomain draw together the viral and cellular membranes. We found stem-derived peptides from dengue virus (DV) bind stem-less E trimer and mimic the stem-reconfiguration step in the fusion pathway. We adapted this experiment as a high-throughput screen for small molecules that block peptide binding and thus may inhibit viral entry. A compound identified in this screen, 1662G07, and a number of its analogs reversibly inhibit DV infectivity. They do so by binding the prefusion, dimeric E on the virion surface, before adsorption to a cell. They also block viral fusion with liposomes. Structure-activity relationship studies have led to analogs with submicromolar IC₉₀s against DV2, and certain analogs are active against DV serotypes 1,2, and 4. The compounds do not inhibit the closely related Kunjin virus. We propose that they bind in a previously identified, E-protein pocket, exposed on the virion surface and although this pocket is closed in the postfusion trimer, its mouth is fully accessible. Examination of the E-trimer coordinates (PDB 1OK8) shows that conformational fluctuations around the hinge could open the pocket without dissociating the trimer or otherwise generating molecular collisions. We propose that compounds such as 1662G07 trap the sE trimer in a "pocket-open" state, which has lost affinity for the stem peptide and cannot support the final "zipping up" of the stem.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
