

Regulation of mat responses by a differentiation MAPK pathway in Saccharomyces cerevisiae
- PMID: 22496730
- PMCID: PMC3319557
- DOI: 10.1371/journal.pone.0032294
Regulation of mat responses by a differentiation MAPK pathway in Saccharomyces cerevisiae
Abstract
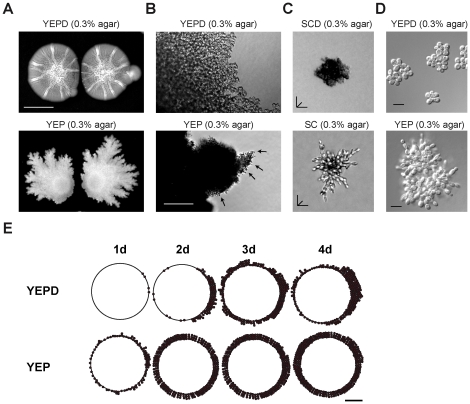
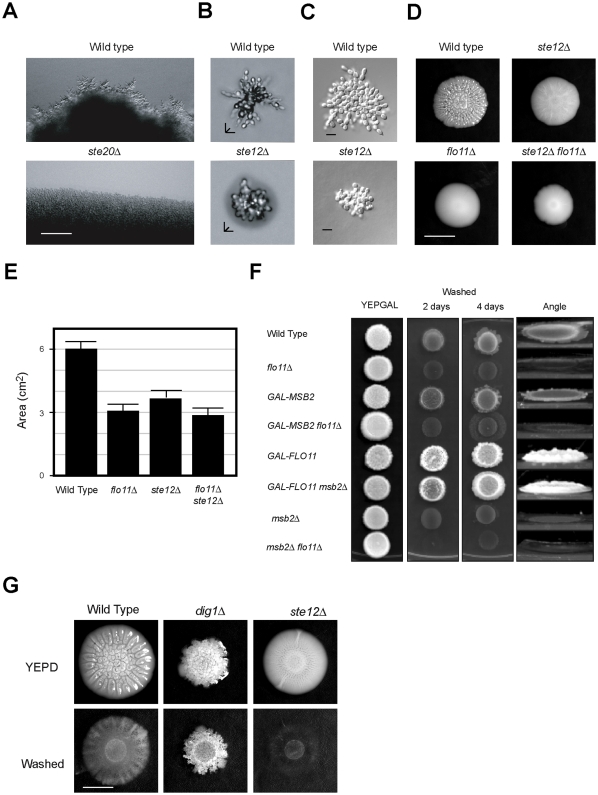
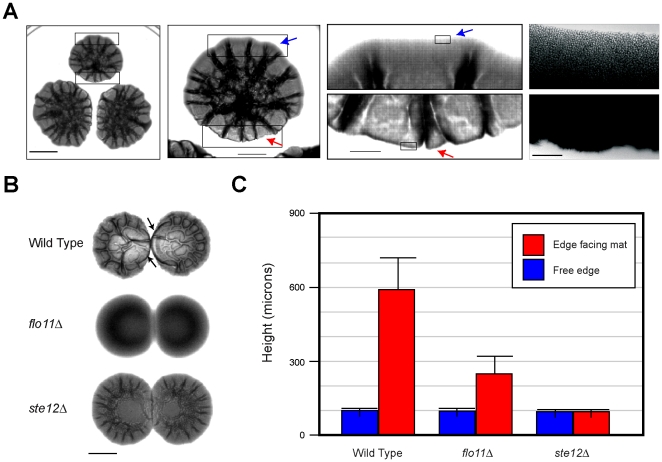
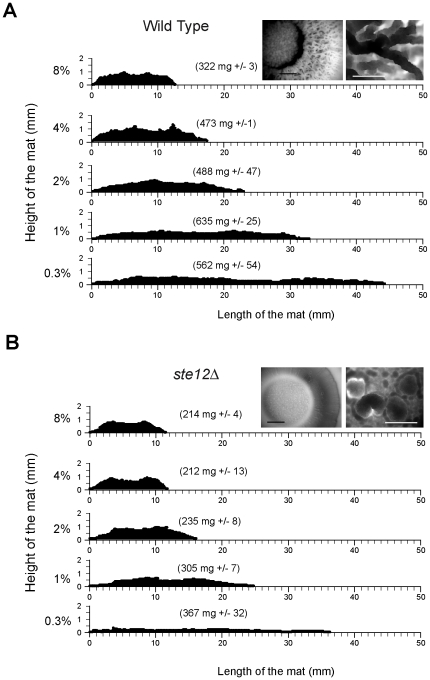
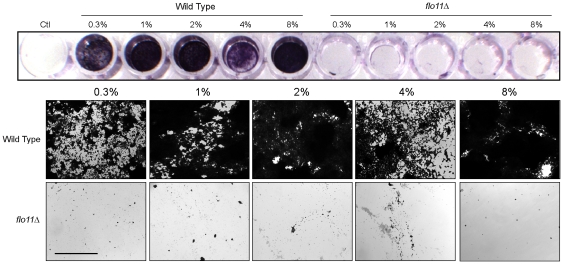
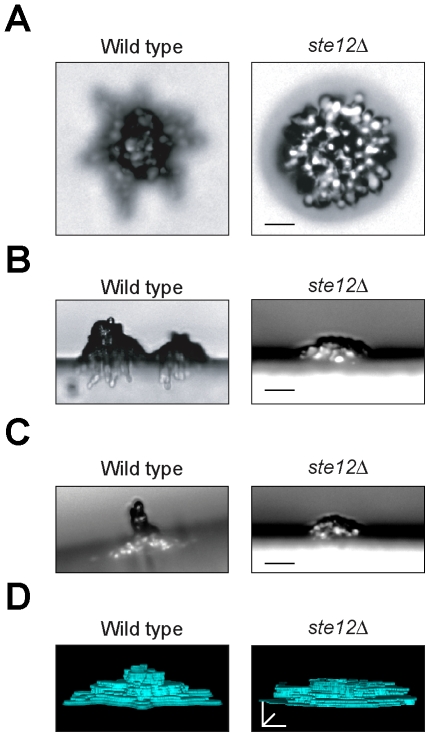
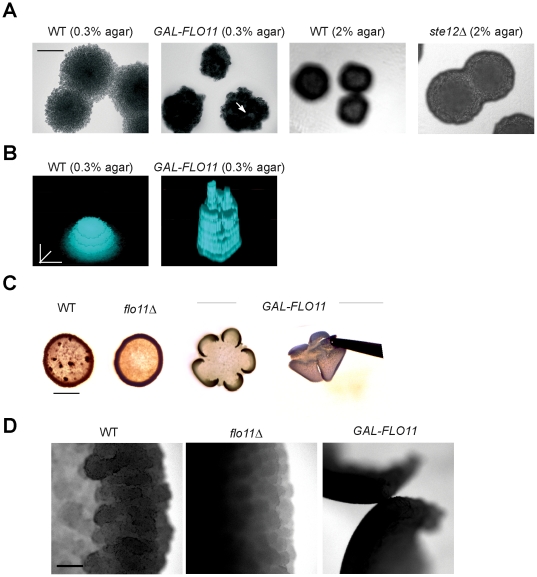
Fungal species exhibit diverse behaviors when presented with extracellular challenges. Pathogenic fungi can undergo cell differentiation and biofilm formation in response to fluctuating nutrient levels, and these responses are required for virulence. In the model fungal eukaryote Saccharomyces cerevisiae, nutrient limitation induces filamentous growth and biofilm/mat formation. Both responses require the same signal transduction (MAPK) pathway and the same cell adhesion molecule (Flo11) but have been studied under different conditions. We found that filamentous growth and mat formation are aspects of a related response that is regulated by the MAPK pathway. Cells in yeast-form mats differentiated into pseudohyphae in response to nutrient limitation. The MAPK pathway regulated mat expansion (in the plane of the XY-axis) and substrate invasion (downward in the plane of the Z-axis), which optimized the mat's response to extracellular nutrient levels. The MAPK pathway also regulated an upward growth pattern (in the plane of the Z-axis) in response to nutrient limitation and changes in surface rigidity. Upward growth allowed for another level of mat responsiveness and resembled a type of colonial chemorepulsion. Together our results show that signaling pathways play critical roles in regulating social behaviors in which fungal cells participate. Signaling pathways may regulate similar processes in pathogens, whose highly nuanced responses are required for virulence.
Conflict of interest statement
Figures

References
-
- Costerton JW, Lewandowski Z, Caldwell DE, Korber DR, Lappin-Scott HM. Microbial biofilms. Annu Rev Microbiol. 1995;49:711–745. - PubMed
-
- Beauvais A, Loussert C, Prevost MC, Verstrepen K, Latge JP. Characterization of a biofilm-like extracellular matrix in FLO1-expressing Saccharomyces cerevisiae cells. FEMS Yeast Res. 2009;9:411–419. - PubMed
-
- Parsek MR, Greenberg EP. Sociomicrobiology: the connections between quorum sensing and biofilms. Trends Microbiol. 2005;13:27–33. - PubMed
-
- Chandra J, Zhou G, Ghannoum MA. Fungal biofilms and antimycotics. Curr Drug Targets. 2005;6:887–894. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
