

F-actin-dependent regulation of NESH dynamics in rat hippocampal neurons
- PMID: 22496823
- PMCID: PMC3319579
- DOI: 10.1371/journal.pone.0034514
F-actin-dependent regulation of NESH dynamics in rat hippocampal neurons
Abstract
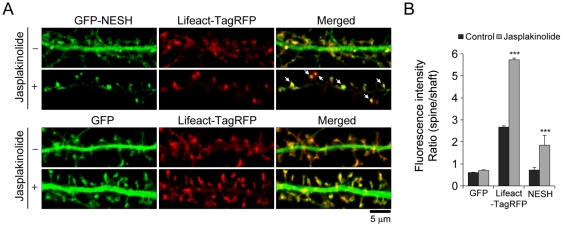
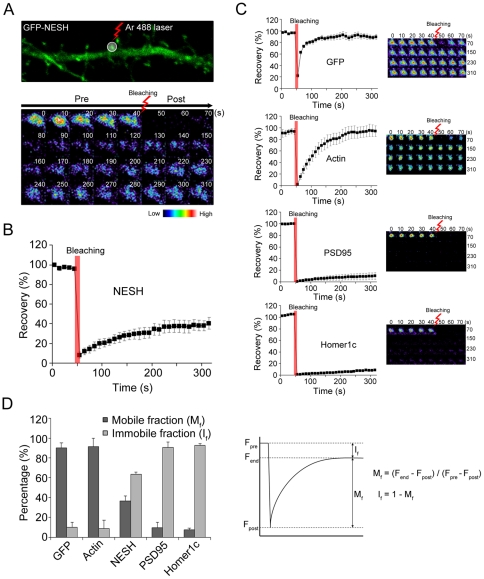
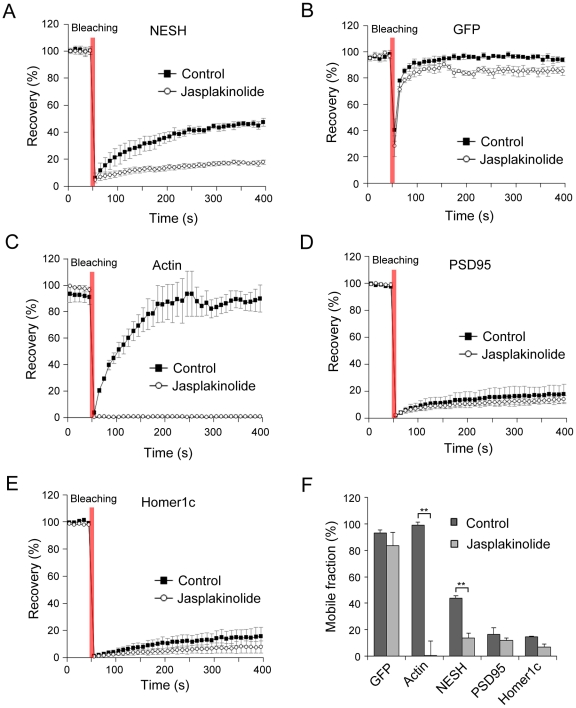
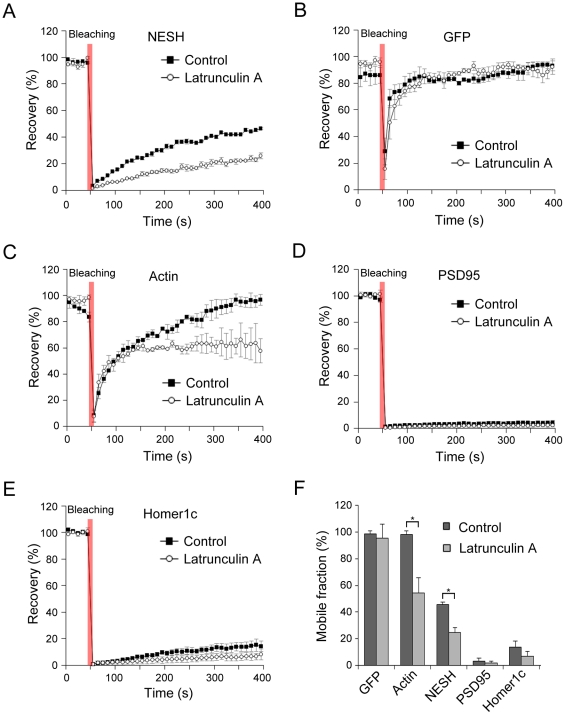
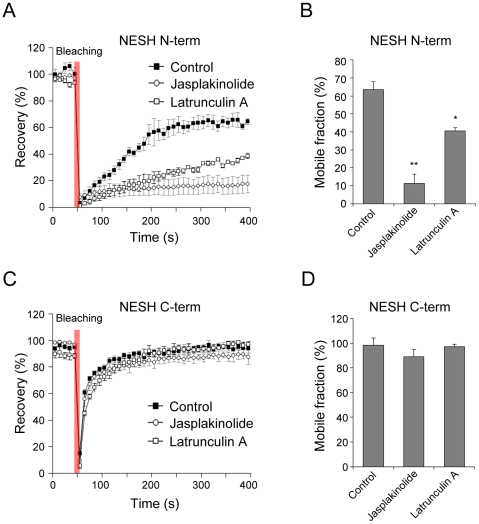
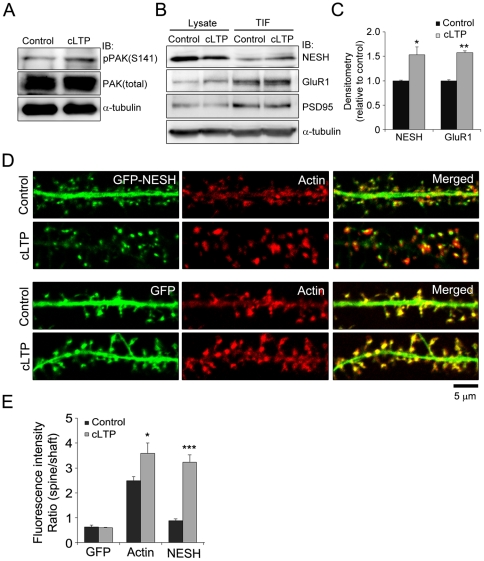
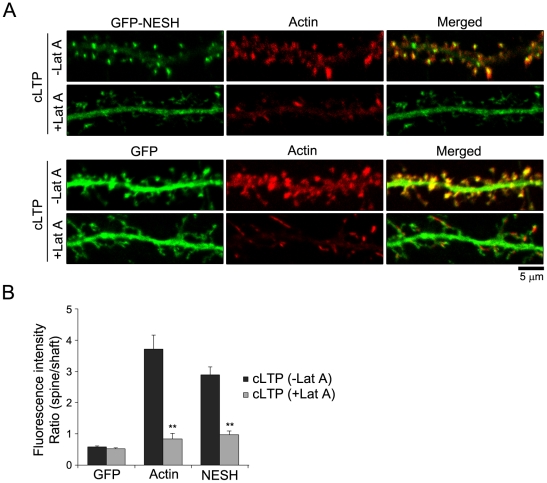
Synaptic plasticity is an important feature of neurons essential for learning and memory. Postsynaptic organization and composition are dynamically remodeled in response to diverse synaptic inputs during synaptic plasticity. During this process, the dynamics and localization of postsynaptic proteins are also precisely regulated. NESH/Abi-3 is a member of the Abl interactor (Abi) protein family. Overexpression of NESH is associated with reduced cell motility and tumor metastasis. Strong evidence of a close relationship between NESH and the actin cytoskeleton has been documented. Although earlier studies have shown that NESH is prominently expressed in the brain, its function and characteristics are yet to be established. Data from the present investigation suggest that synaptic localization of NESH in hippocampal neurons is regulated in an F-actin-dependent manner. The dynamic fraction of NESH in the dendritic spine was analyzed using FRAP (fluorescence recovery after photobleaching). Interestingly, F-actin stabilization and disruption significantly affected the mobile fraction of NESH, possibly through altered interactions of NESH with the F-actin. In addition, NESH was synaptically targeted from the dendritic shaft to spine after induction of chemical LTP (long-term potentiation) and the translocation was dependent on F-actin. Our data collectively support the significance of the F-actin cytoskeleton in synaptic targeting of NESH as well as its dynamics.
Conflict of interest statement
Figures
References
-
- Sheng M, Kim MJ. Postsynaptic signaling and plasticity mechanisms. Science. 2002;298:776–780. - PubMed
-
- McGee AW, Bredt DS. Assembly and plasticity of the glutamatergic postsynaptic specialization. Current opinion in neurobiology. 2003;13:111–118. - PubMed
-
- Sheng M, Hoogenraad CC. The postsynaptic architecture of excitatory synapses: a more quantitative view. Annual review of biochemistry. 2007;76:823–847. - PubMed
-
- Renner M, Specht CG, Triller A. Molecular dynamics of postsynaptic receptors and scaffold proteins. Current opinion in neurobiology. 2008;18:532–540. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
