

Cleavage of PGRP-LC receptor in the Drosophila IMD pathway in response to live bacterial infection in S2 cells
- PMID: 22496930
- PMCID: PMC3323661
- DOI: 10.4161/self.17882
Cleavage of PGRP-LC receptor in the Drosophila IMD pathway in response to live bacterial infection in S2 cells
Abstract
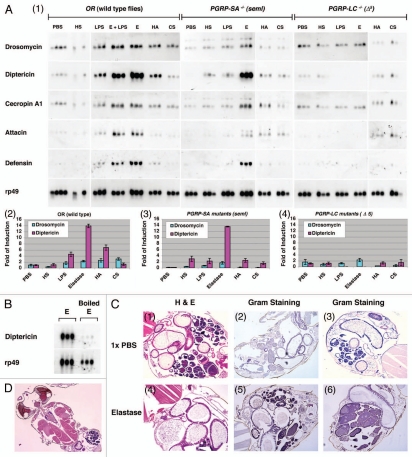
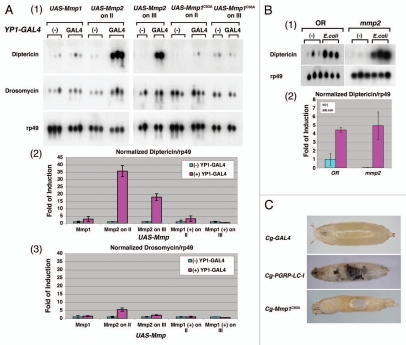
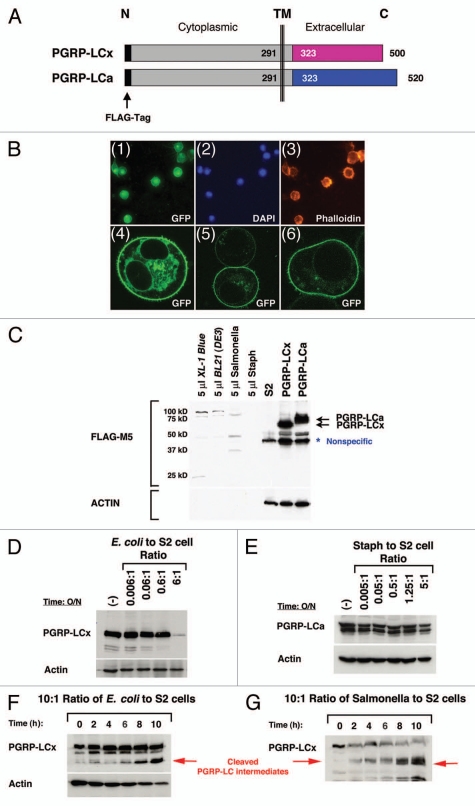
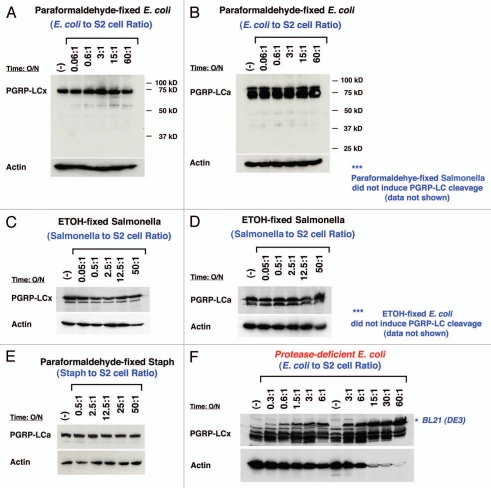
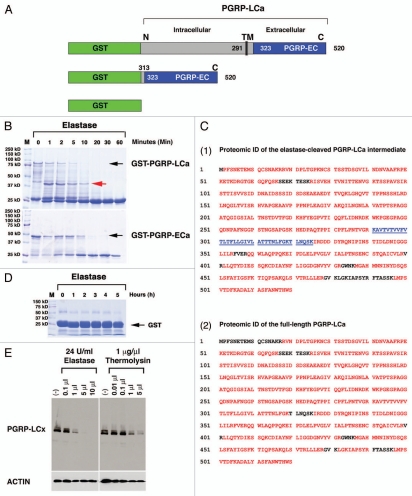
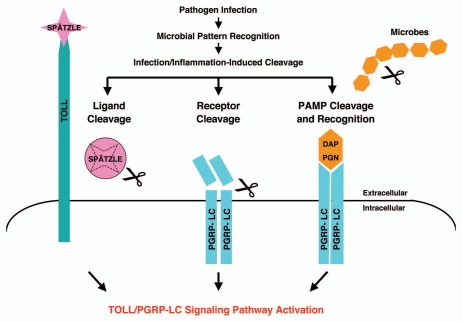
Drosophila responds to Gram-negative bacterial infection by activating the immune deficiency (IMD) pathway, leading to production of antimicrobial peptides (AMPs). As a receptor for the IMD pathway, peptidoglycan-recognition protein (PGRP), PGRP-LC is known to recognize and bind monomeric peptidoglycan (DAP-type PGN) through its PGRP ectodomain and in turn activate the IMD pathway. The questions remain how PGRP-LC is activated in response to pathogen infection to initiate the IMD signal transduction in Drosophila. Here we present evidence to show that proteases such as elastase and Mmp2 can also activate the IMD pathway but not the TOLL pathway. The elastase-dependent IMD activation requires the receptor PGRP-LC. Importantly, we find that live Salmonella/E. coli infection modulates PGRP-LC expression/receptor integrity and activates the IMD pathway while dead Salmonella/E. coli or protease-deficient E. coli do neither. Our results suggest an interesting possibility that Gram-negative pathogen infection may be partially monitored through the structural integrity of the receptor PGRP-LC via an infection-induced enzyme-based cleavage-mediated activation mechanism.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous