

Therapeutic potential of HIV protease-activable CASP3
- PMID: 22496955
- PMCID: PMC3323887
- DOI: 10.1038/srep00359
Therapeutic potential of HIV protease-activable CASP3
Abstract
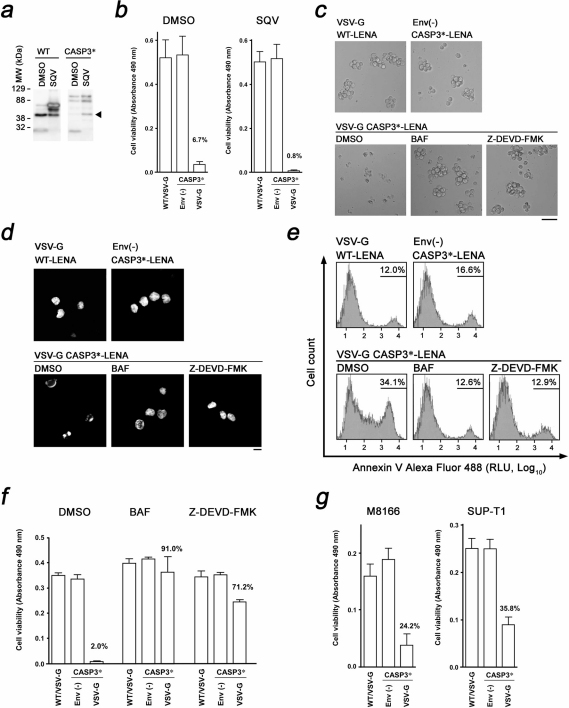
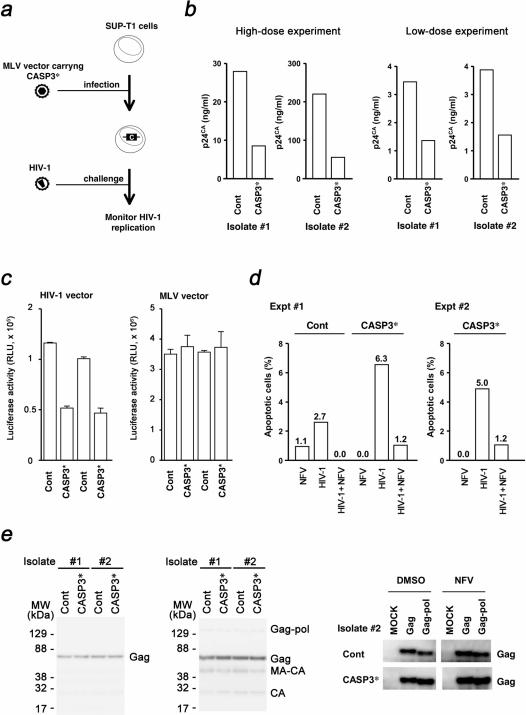
Development of a therapeutic application of CASP3/caspase 3/CPP32, an executor of apoptosis, has been challenging because regulation of its activation is complicated. This study aimed to inhibit cancer cell growth and human immunodeficiency virus type 1 (HIV-1) propagation through a CASP3 mutant, CASP3*, activable by HIV-1-encoded aspartate protease. Active CASP3* was delivered to leukemic cells using a protein transduction vehicle, the lentivirus-like nanoparticle (LENA), which should contain thousands of CASP3*-Gag protein molecules and release the activated CASP3* into the target cell cytoplasm. CASP3*-LENA induced apoptosis in various types of leukemic cells. In addition to being effective against leukemic cells, constitutive expression of CASP3* restricted HIV-1 propagation in SUP-T1 cells. The attenuation of HIV-1 replication in SUP-T1/CASP3* cells was attributed to the elimination of HIV-1-infected cells by apoptosis. These data suggest that CASP3* has therapeutic potential against both lymphoid malignancies and HIV-1 infection.
Figures

References
-
- Boatright K. M. & Salvesen G. S. Mechanisms of caspase activation. Curr Opin Cell Biol. 15, 725–31 (2003). - PubMed
-
- Yamabe K. et al. Cancer gene therapy using a pro-apoptotic gene, caspase-3. Gene Ther. 6, 1952–9 (1999). - PubMed
-
- Cam L., Boucquey A., Coulomb-L'hermine A., Weber A. & Horellou P. Gene transfer of constitutively active caspase-3 induces apoptosis in a human hepatoma cell line. J Gene Med. 7, 30–8 (2005). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
