

Proteolytic regulation of alginate overproduction in Pseudomonas aeruginosa
- PMID: 22497280
- PMCID: PMC3345095
- DOI: 10.1111/j.1365-2958.2012.08049.x
Proteolytic regulation of alginate overproduction in Pseudomonas aeruginosa
Abstract
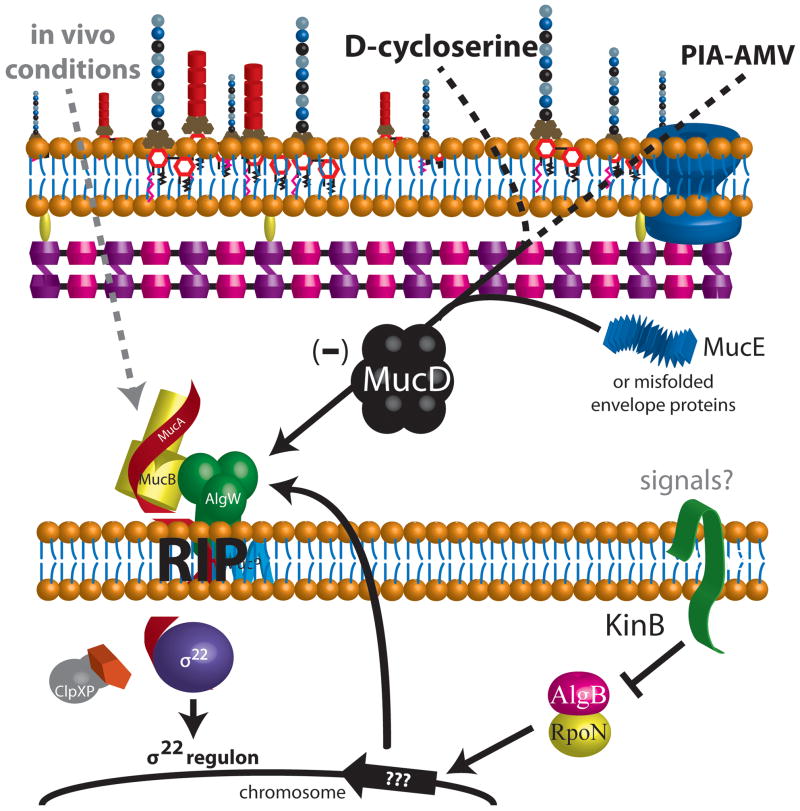
Pseudomonas aeruginosa, a Gram-negative bacterium, is a significant opportunistic pathogen associated with skin and soft tissue infections, nosocomial pneumonia and sepsis. In addition, it can chronically colonize the lungs of cystic fibrosis (CF) patients. Overproduction of the exopolysaccharide called alginate provides P. aeruginosa with a selective advantage and facilitates survival in the CF lung. The in vitro phenotype of alginate overproduction observed on solid culture media is referred to as mucoid. Expression of the alginate machinery and biosynthetic enzymes are controlled by the extracytoplasmic sigma factor, σ(22) (AlgU/T). The key negative regulator of both σ(22) activity and the mucoid phenotype is the cognate anti-sigma factor MucA. MucA sequesters σ(22) to the inner membrane inhibiting the sigma factor's transcriptional activity. The well-studied mechanism for transition to the mucoid phenotype is mutation of mucA, leading to loss of MucA function and therefore activation of σ(22) . Recently, regulated intramembrane proteolysis (RIP) has been recognized as a mechanism whereby proteolysis of the anti-sigma factor MucA leads to active σ(22) allowing P. aeruginosa to respond to environmental stress conditions by overproduction of alginate. The goal of this review is to illuminate the pathways leading to RIP that have been identified and proposed.
© 2012 Blackwell Publishing Ltd.
Figures



References
-
- Ades SE. Regulation by destruction: design of the sigmaE envelope stress response. Curr Opin Microbiol. 2008;11:535–540. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
