

Axonal mRNA localization and local protein synthesis in nervous system assembly, maintenance and repair
- PMID: 22498899
- PMCID: PMC3682205
- DOI: 10.1038/nrn3210
Axonal mRNA localization and local protein synthesis in nervous system assembly, maintenance and repair
Erratum in
- Nat Rev Neurosci. 2012 Aug;13(8):597
Abstract
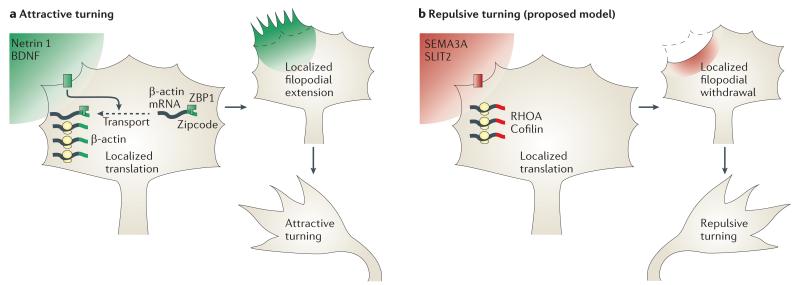
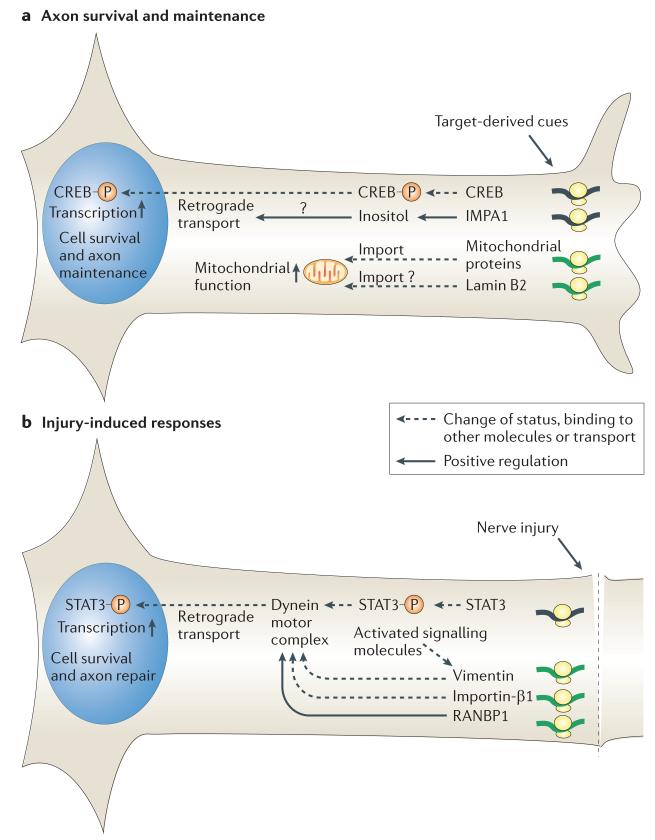
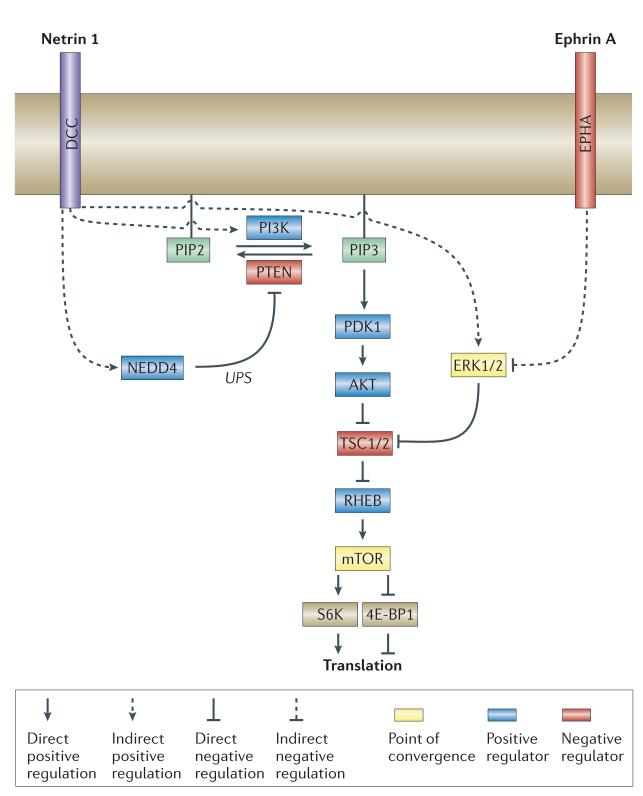
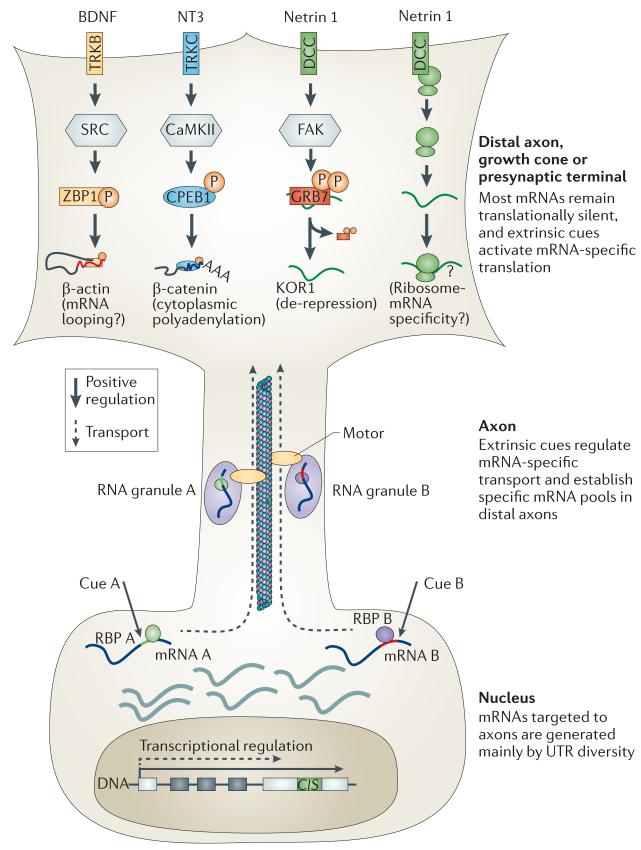
mRNAs can be targeted to specific neuronal subcellular domains, which enables rapid changes in the local proteome through local translation. This mRNA-based mechanism links extrinsic signals to spatially restricted cellular responses and can mediate stimulus-driven adaptive responses such as dendritic plasticity. Local mRNA translation also occurs in growing axons where it can mediate directional responses to guidance signals. Recent profiling studies have revealed that both growing and mature axons possess surprisingly complex and dynamic transcriptomes, thereby suggesting that axonal mRNA localization is highly regulated and has a role in a broad range of processes, a view that is increasingly being supported by new experimental evidence. Here, we review current knowledge on the roles and regulatory mechanisms of axonal mRNA translation and discuss emerging links to axon guidance, survival, regeneration and neurological disorders.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
