

NLRP12 suppresses colon inflammation and tumorigenesis through the negative regulation of noncanonical NF-κB signaling
- PMID: 22503542
- PMCID: PMC3658309
- DOI: 10.1016/j.immuni.2012.03.012
NLRP12 suppresses colon inflammation and tumorigenesis through the negative regulation of noncanonical NF-κB signaling
Abstract
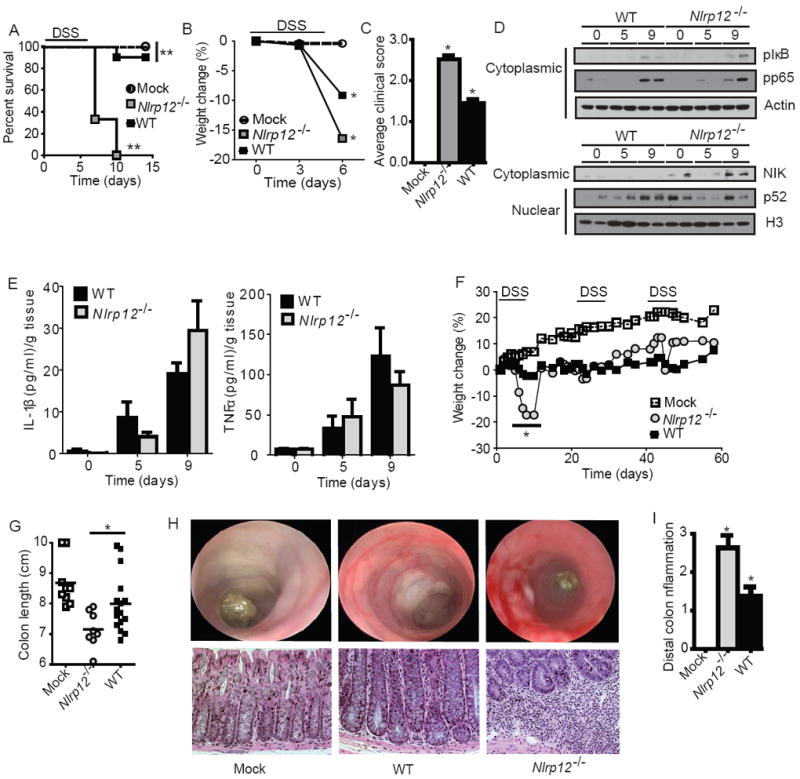
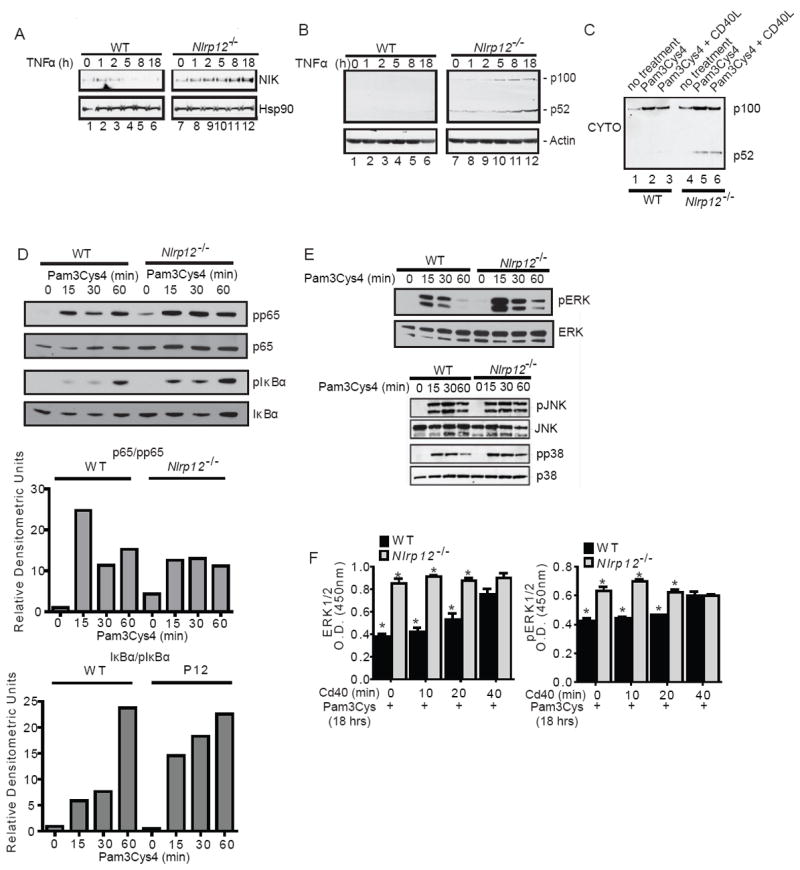
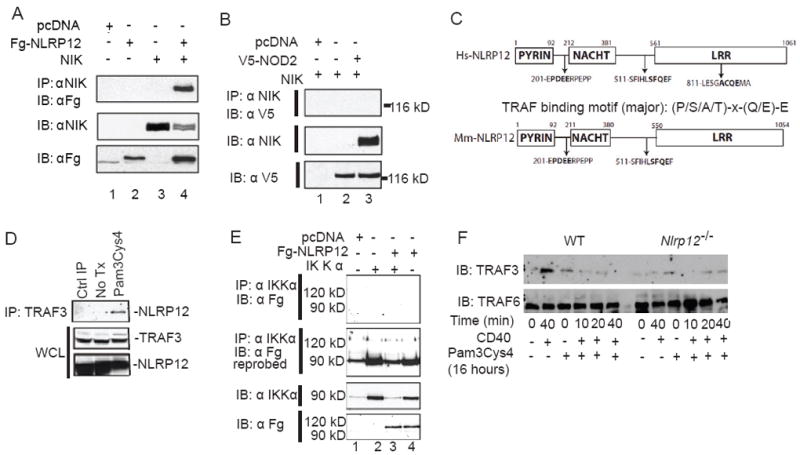
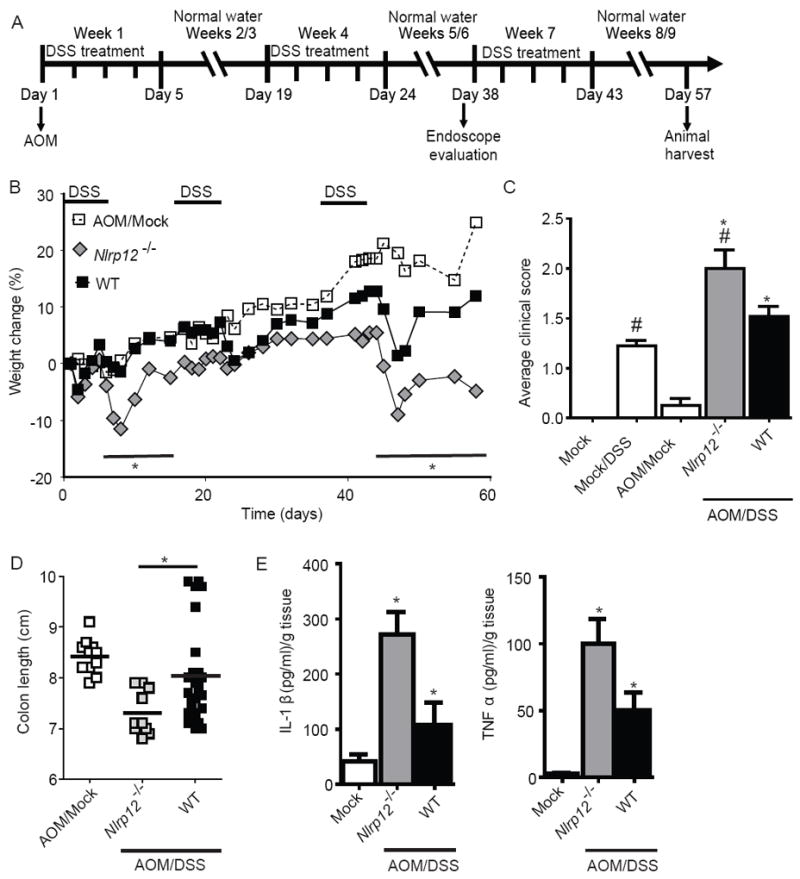
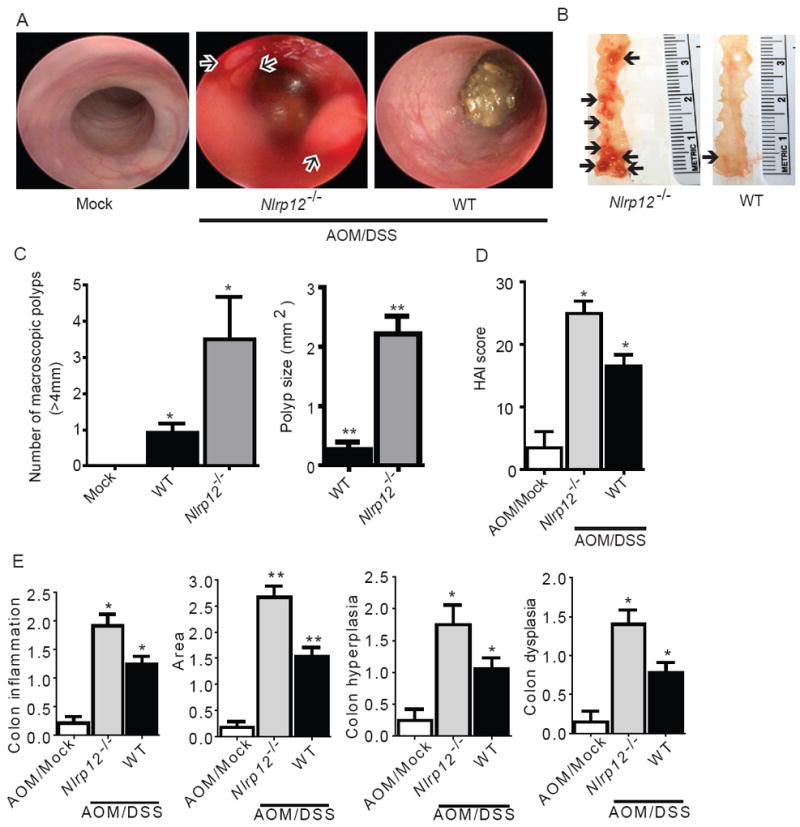
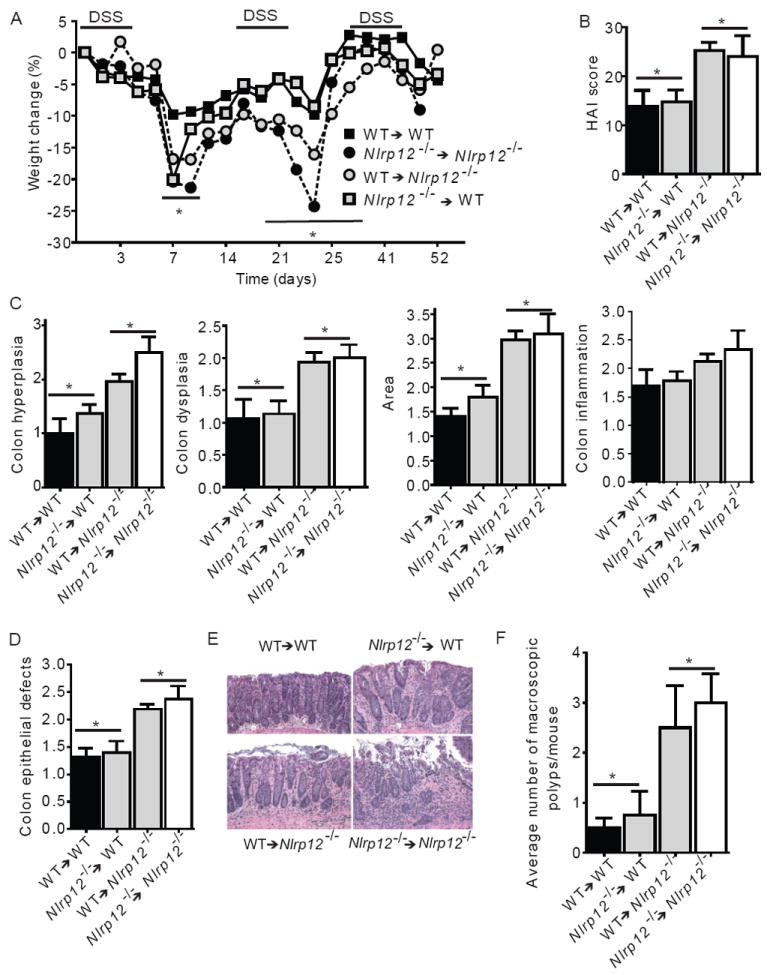
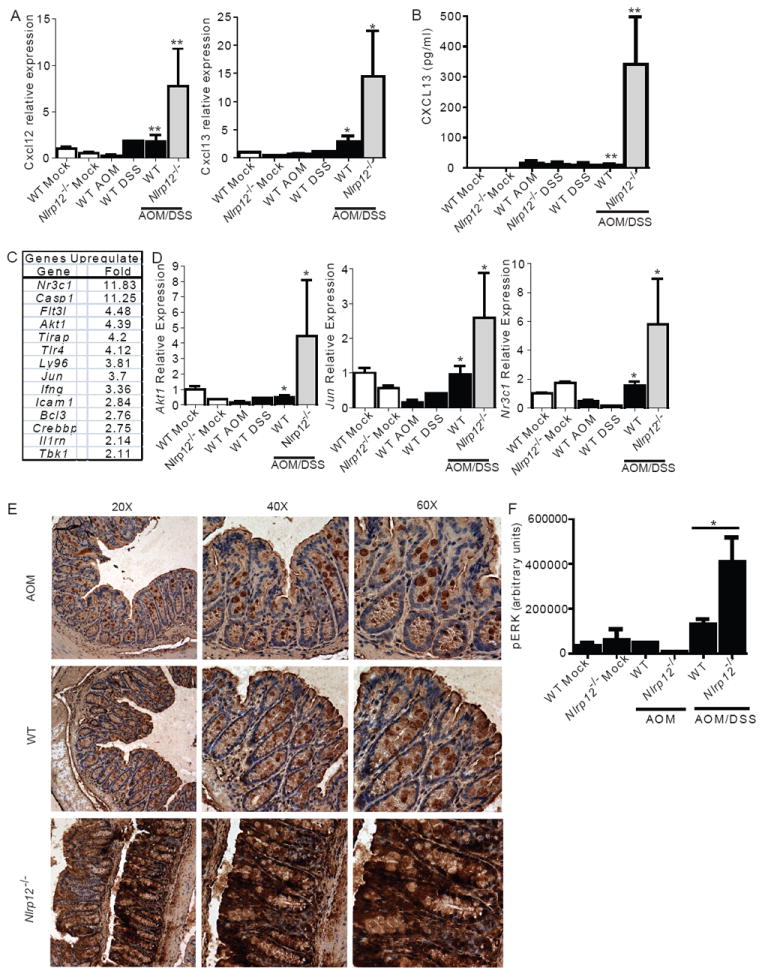
In vitro data suggest that a subgroup of NLR proteins, including NLRP12, inhibits the transcription factor NF-κB, although physiologic and disease-relevant evidence is largely missing. Dysregulated NF-κB activity is associated with colonic inflammation and cancer, and we found Nlrp12(-/-) mice were highly susceptible to colitis and colitis-associated colon cancer. Polyps isolated from Nlrp12(-/-) mice showed elevated noncanonical NF-κB activation and increased expression of target genes that were associated with cancer, including Cxcl13 and Cxcl12. NLRP12 negatively regulated ERK and AKT signaling pathways in affected tumor tissues. Both hematopoietic- and nonhematopoietic-derived NLRP12 contributed to inflammation, but the latter dominantly contributed to tumorigenesis. The noncanonical NF-κB pathway was regulated upon degradation of TRAF3 and activation of NIK. NLRP12 interacted with both NIK and TRAF3, and Nlrp12(-/-) cells have constitutively elevated NIK, p100 processing to p52 and reduced TRAF3. Thus, NLRP12 is a checkpoint of noncanonical NF-κB, inflammation, and tumorigenesis.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK073338/DK/NIDDK NIH HHS/United States
- F32 DK088417/DK/NIDDK NIH HHS/United States
- R21 CA131645/CA/NCI NIH HHS/United States
- P30 DK034987/DK/NIDDK NIH HHS/United States
- P30 CA016086/CA/NCI NIH HHS/United States
- AI077437/AI/NIAID NIH HHS/United States
- K01 DK092355/DK/NIDDK NIH HHS/United States
- CA156330/CA/NCI NIH HHS/United States
- AR007416/AR/NIAMS NIH HHS/United States
- U19 AI067798/AI/NIAID NIH HHS/United States
- P30DK34987/DK/NIDDK NIH HHS/United States
- R01 CA156330/CA/NCI NIH HHS/United States
- R21CA131645/CA/NCI NIH HHS/United States
- AI067798/AI/NIAID NIH HHS/United States
- K01DK092355/DK/NIDDK NIH HHS/United States
- T32 AR007416/AR/NIAMS NIH HHS/United States
- U19 AI077437/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
