

Adora2b-elicited Per2 stabilization promotes a HIF-dependent metabolic switch crucial for myocardial adaptation to ischemia
- PMID: 22504483
- PMCID: PMC3378044
- DOI: 10.1038/nm.2728
Adora2b-elicited Per2 stabilization promotes a HIF-dependent metabolic switch crucial for myocardial adaptation to ischemia
Abstract
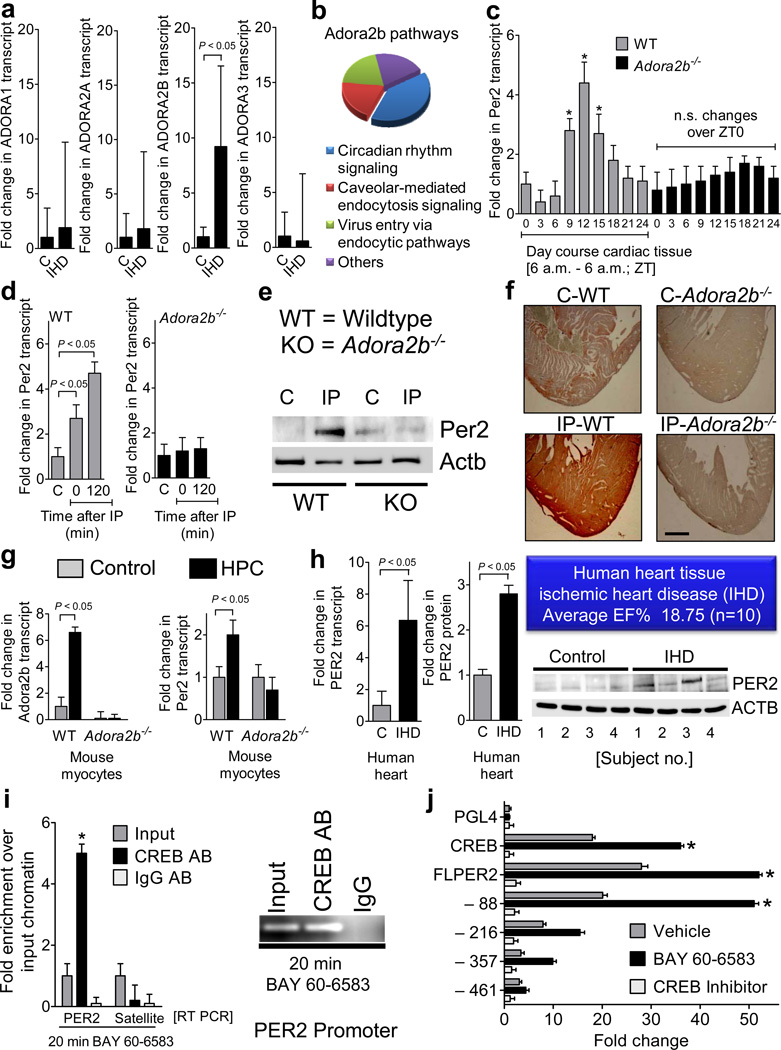
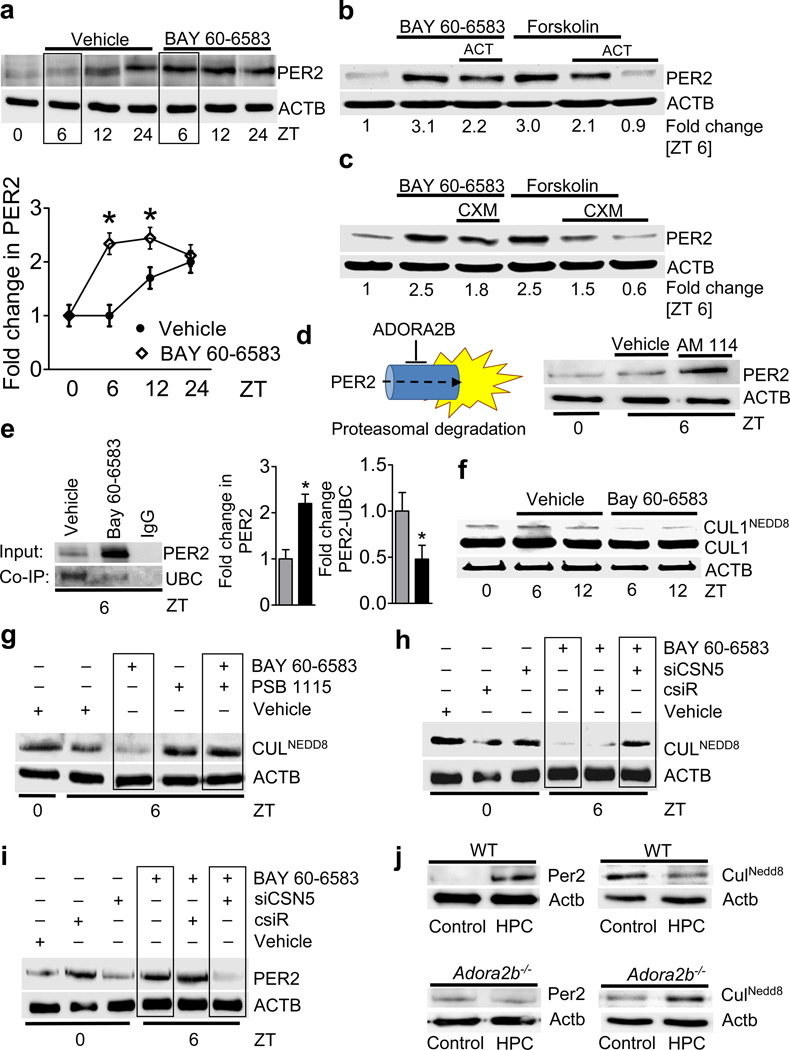
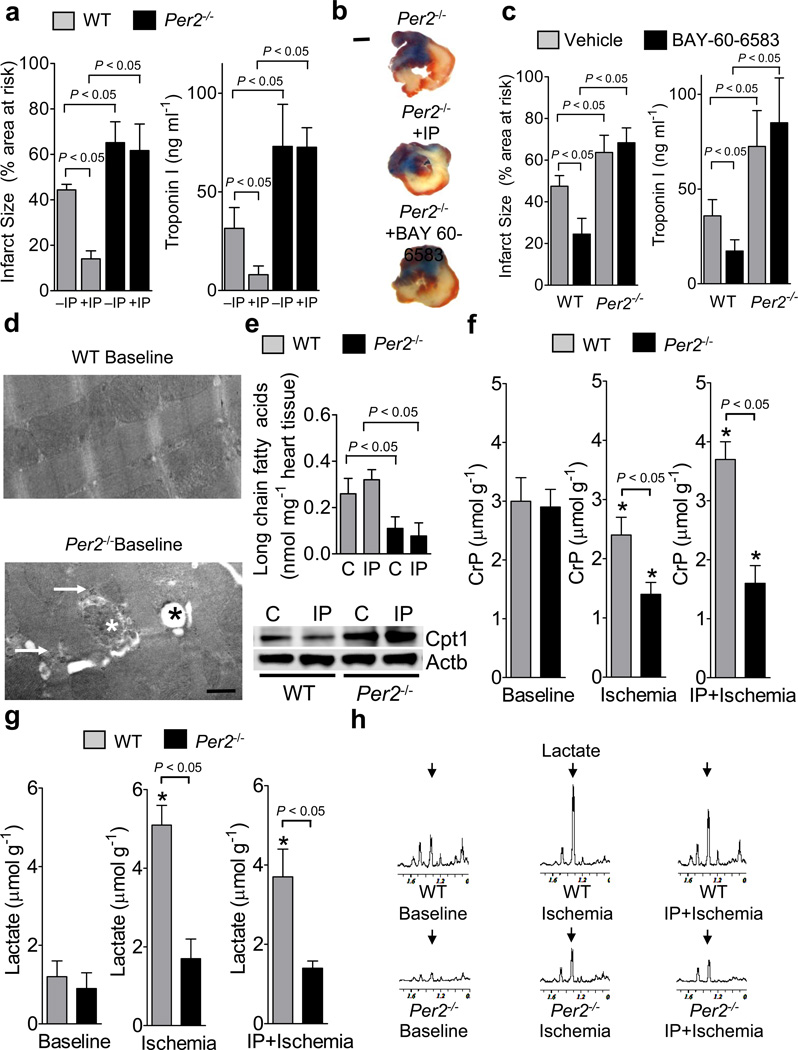
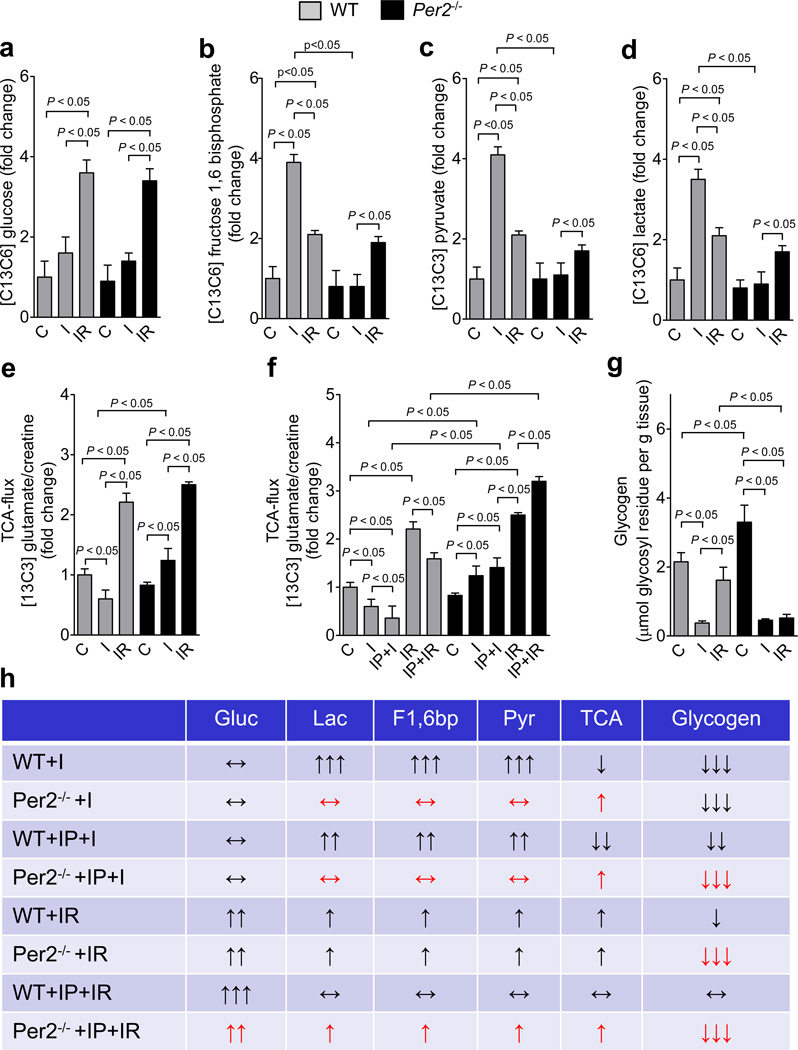
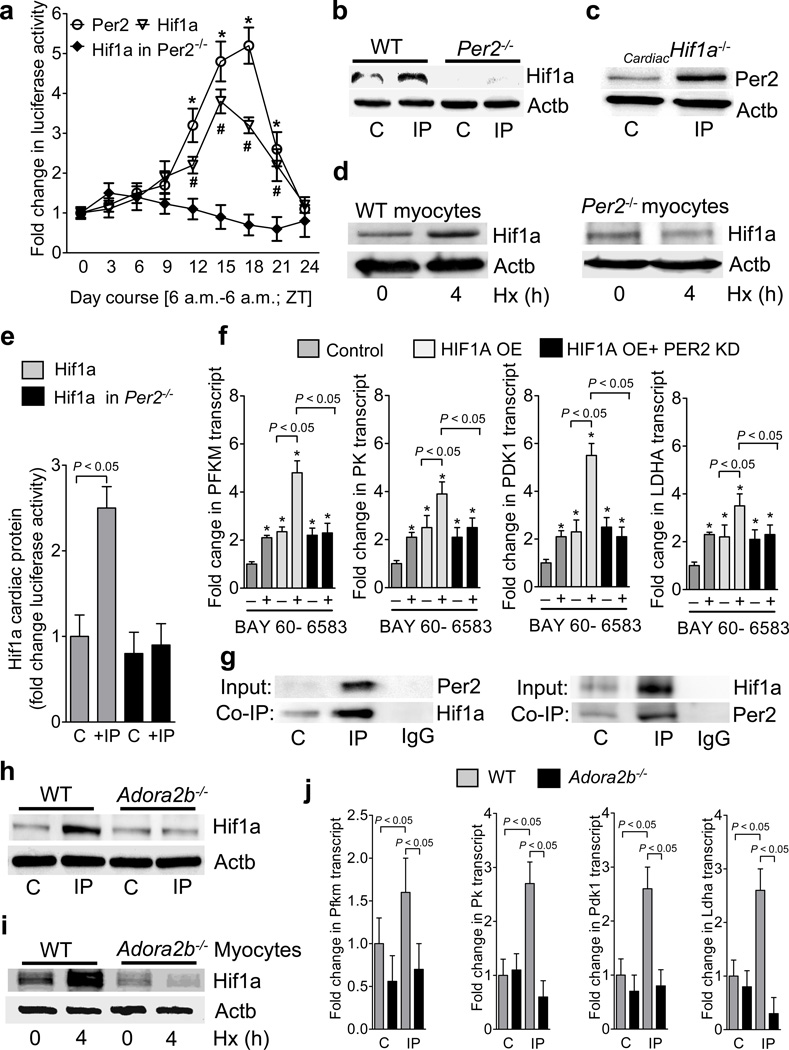
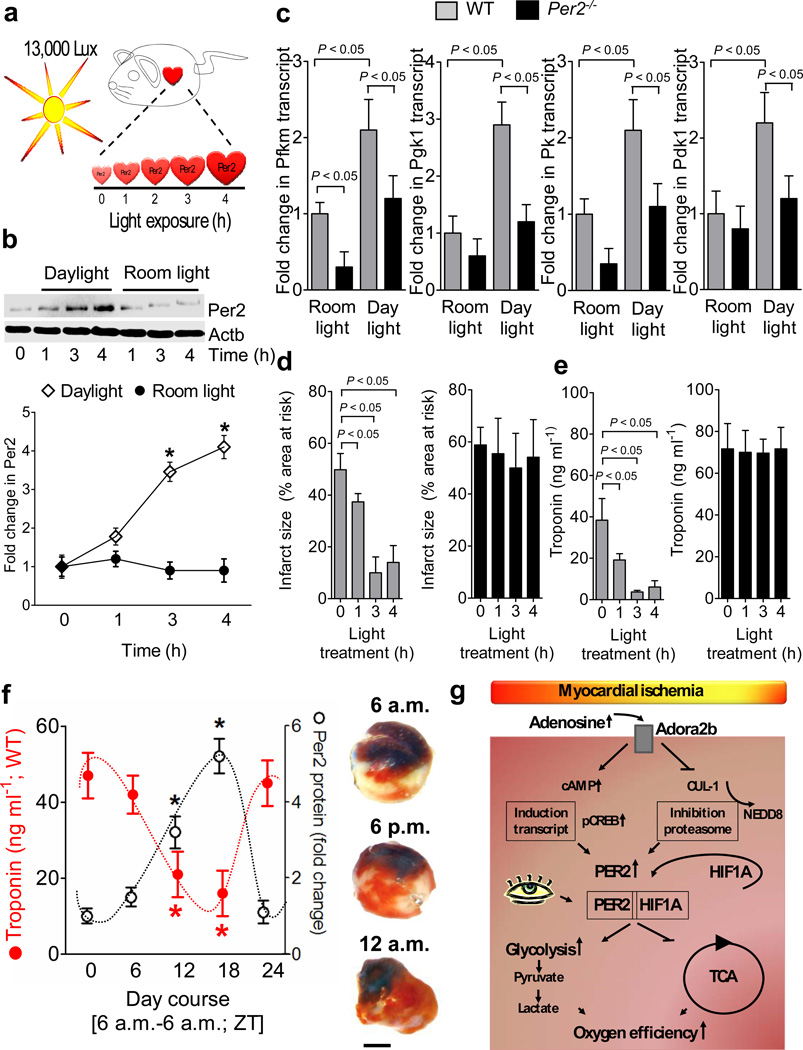
Adenosine signaling has been implicated in cardiac adaptation to limited oxygen availability. In a wide search for adenosine receptor A2b (Adora2b)-elicited cardioadaptive responses, we identified the circadian rhythm protein period 2 (Per2) as an Adora2b target. Adora2b signaling led to Per2 stabilization during myocardial ischemia, and in this setting, Per2(-/-) mice had larger infarct sizes compared to wild-type mice and loss of the cardioprotection conferred by ischemic preconditioning. Metabolic studies uncovered a limited ability of ischemic hearts in Per2(-/-) mice to use carbohydrates for oxygen-efficient glycolysis. This impairment was caused by a failure to stabilize hypoxia-inducible factor-1α (Hif-1α). Moreover, stabilization of Per2 in the heart by exposing mice to intense light resulted in the transcriptional induction of glycolytic enzymes and Per2-dependent cardioprotection from ischemia. Together, these studies identify adenosine-elicited stabilization of Per2 in the control of HIF-dependent cardiac metabolism and ischemia tolerance and implicate Per2 stabilization as a potential new strategy for treating myocardial ischemia.
Figures
Comment in
-
A role for period 2 in cardioprotection.Cell Metab. 2012 Jul 3;16(1):2-4. doi: 10.1016/j.cmet.2012.06.008. Cell Metab. 2012. PMID: 22768832
-
(Almost) everything is illuminated: adenosine shines a light on cardioprotection.Circ Res. 2012 Sep 28;111(8):965-6. doi: 10.1161/CIRCRESAHA.112.279752. Circ Res. 2012. PMID: 23023508 No abstract available.
References
-
- Sitkovsky MV, et al. Physiological control of immune response and inflammatory tissue damage by hypoxia-inducible factors and adenosine A2A receptors. Annual Review of Immunology. 2004;22:657–682. - PubMed
-
- Neubauer S. The failing heart--an engine out of fuel. N Engl J Med. 2007;356:1140–1151. - PubMed
-
- Aragones J, Fraisl P, Baes M, Carmeliet P. Oxygen Sensors at the Crossroad of Metabolism. Cell Metab. 2009;9:11–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01-HL092188/HL/NHLBI NIH HHS/United States
- 1K08HL102267/HL/NHLBI NIH HHS/United States
- R01 DK083385/DK/NIDDK NIH HHS/United States
- K08 HL102267/HL/NHLBI NIH HHS/United States
- R01-HL060569/HL/NHLBI NIH HHS/United States
- R01 HL098294/HL/NHLBI NIH HHS/United States
- R01 HL060569/HL/NHLBI NIH HHS/United States
- R01-HL098294/HL/NHLBI NIH HHS/United States
- R01 HL092188/HL/NHLBI NIH HHS/United States
- R37 DK050189/DK/NIDDK NIH HHS/United States
- R01 DK095491/DK/NIDDK NIH HHS/United States
- R01-DK083385/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
