

Structure of the c(10) ring of the yeast mitochondrial ATP synthase in the open conformation
- PMID: 22504883
- PMCID: PMC3343227
- DOI: 10.1038/nsmb.2284
Structure of the c(10) ring of the yeast mitochondrial ATP synthase in the open conformation
Abstract
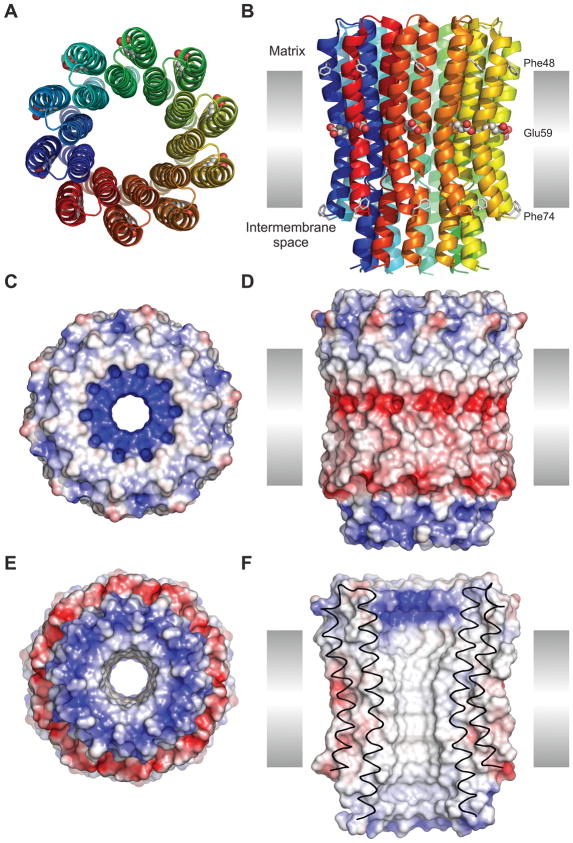
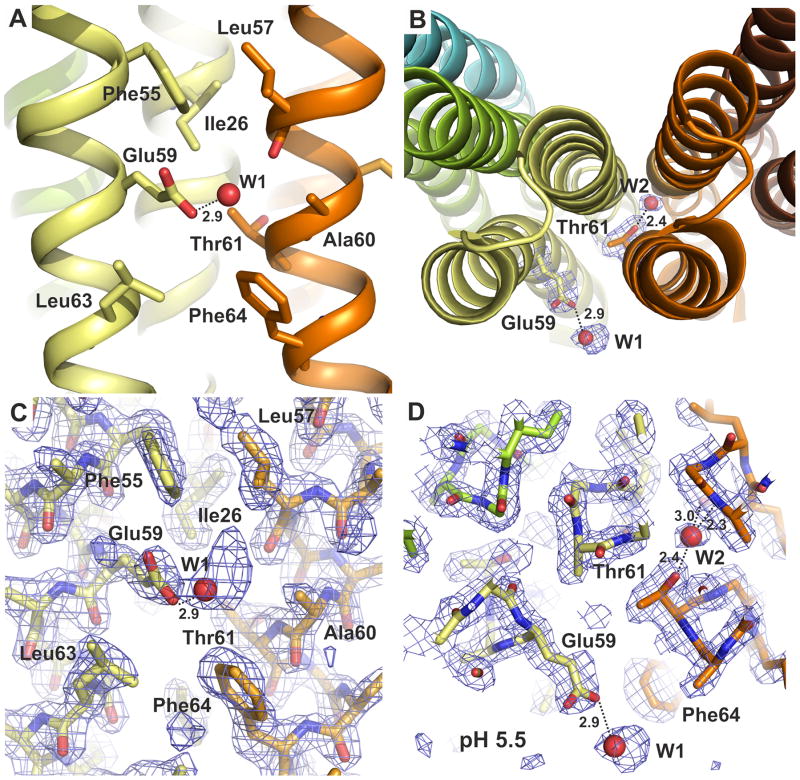
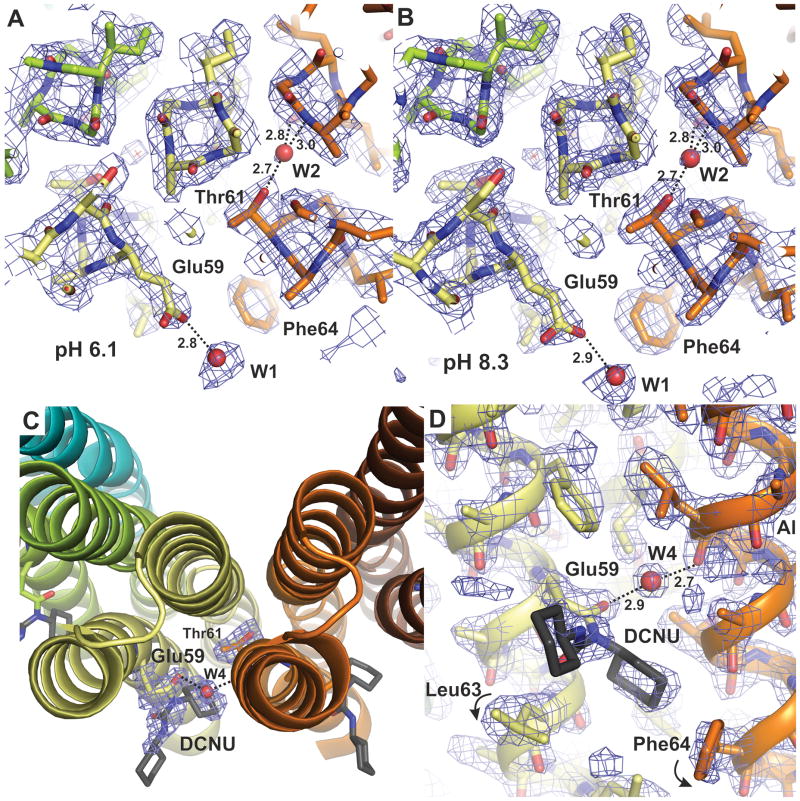
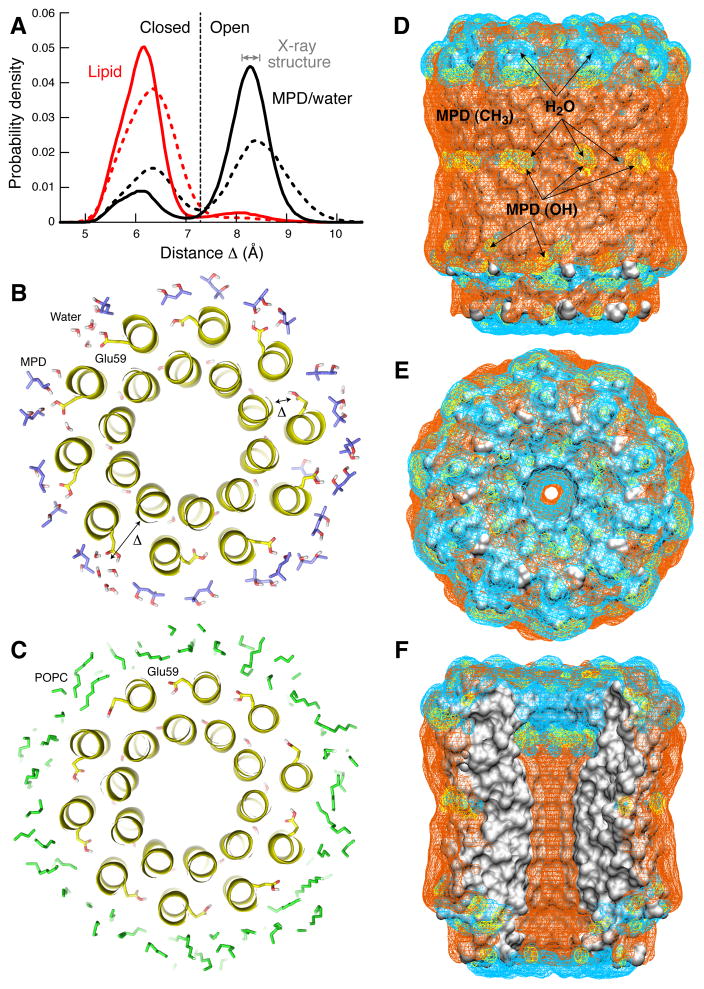
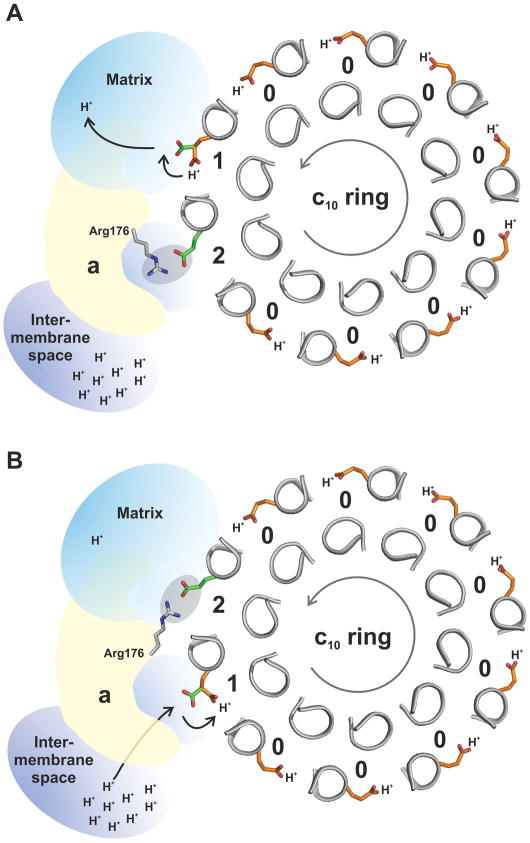
The proton pore of the F(1)F(o) ATP synthase consists of a ring of c subunits, which rotates, driven by downhill proton diffusion across the membrane. An essential carboxylate side chain in each subunit provides a proton-binding site. In all the structures of c-rings reported to date, these sites are in a closed, ion-locked state. Structures are here presented of the c(10) ring from Saccharomyces cerevisiae determined at pH 8.3, 6.1 and 5.5, at resolutions of 2.0 Å, 2.5 Å and 2.0 Å, respectively. The overall structure of this mitochondrial c-ring is similar to known homologs, except that the essential carboxylate, Glu59, adopts an open extended conformation. Molecular dynamics simulations reveal that opening of the essential carboxylate is a consequence of the amphiphilic nature of the crystallization buffer. We propose that this new structure represents the functionally open form of the c subunit, which facilitates proton loading and release.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Collinson IR, Skehel JM, Fearnley IM, Runswick MJ, Walker JE. The F1F0-ATPase complex from bovine heart mitochondria: The molar ratio of the subunits in the stalk region linking the F1 and F0 domains. Biochemistry. 1996;35:12640–12646. - PubMed
-
- Collinson IR, et al. Fo membrane domain of ATP synthase from bovine heart mitochondria: purification, subunit composition, and reconstitution with F1-ATPase. Biochemistry. 1994;33:7971–7978. - PubMed
-
- Stock D, Leslie AGW, Walker JE. Molecular architecture of the rotary motor in ATP synthase. Science. 1999;286:1700–1705. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
