

Argonaute identity defines the length of mature mammalian microRNAs
- PMID: 22505576
- PMCID: PMC3413106
- DOI: 10.1093/nar/gks293
Argonaute identity defines the length of mature mammalian microRNAs
Abstract
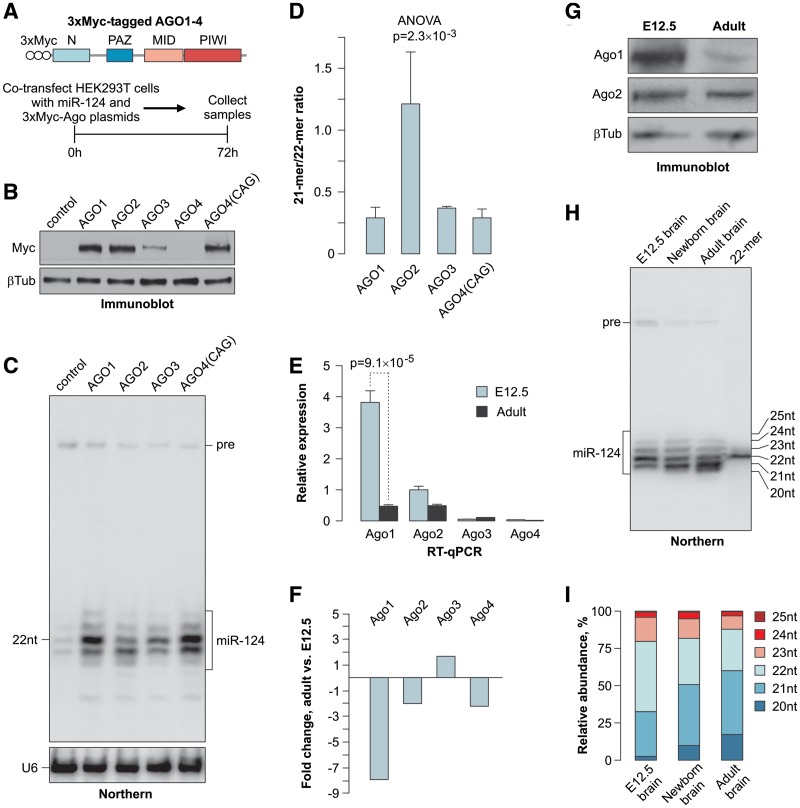
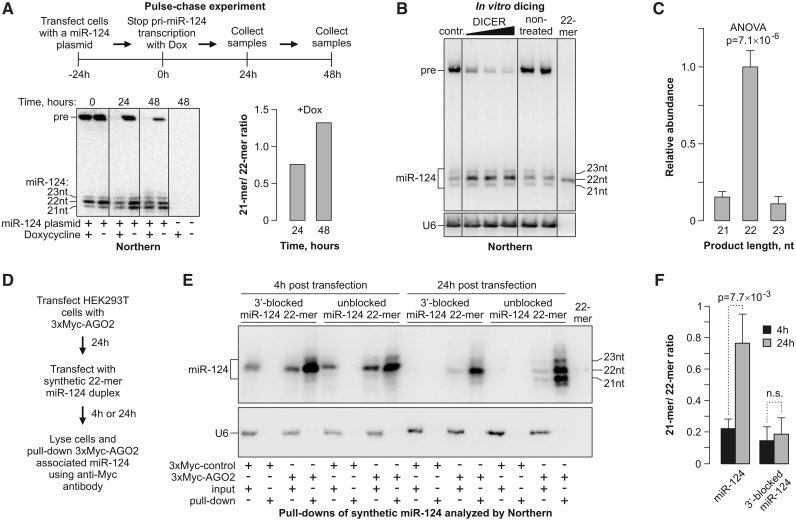
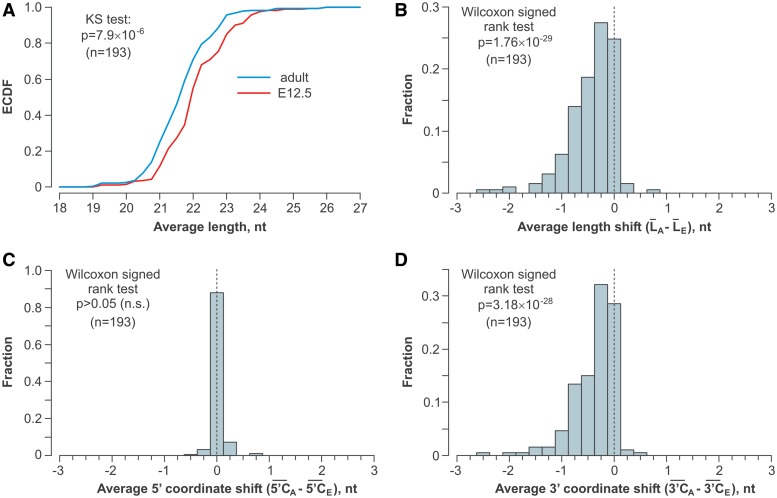
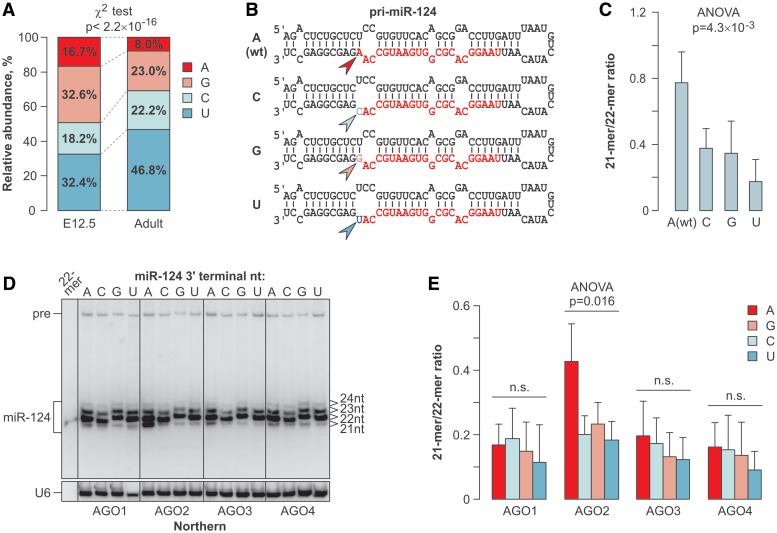
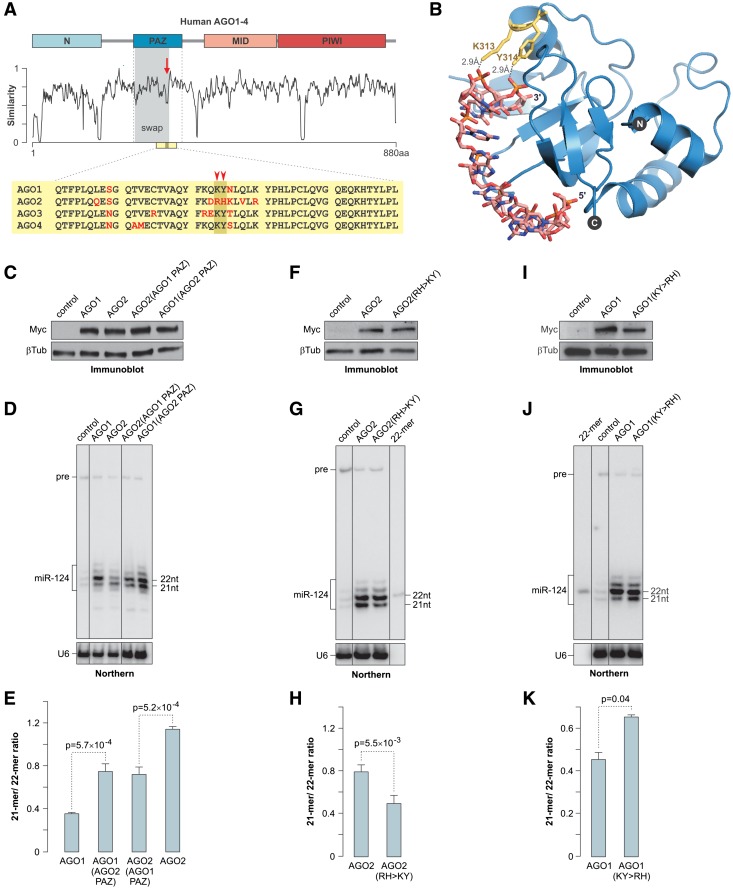
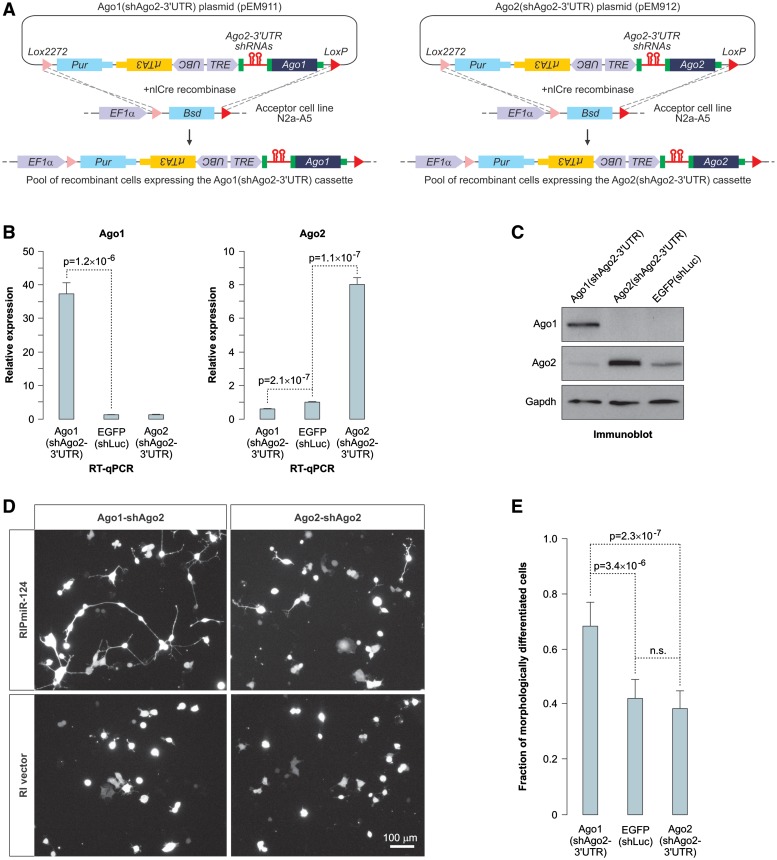
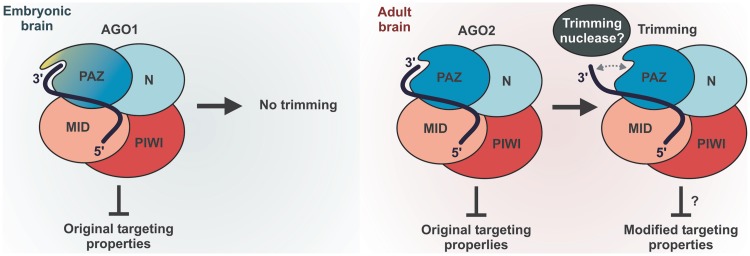
MicroRNAs (miRNAs) are 19- to 25-nt-long non-coding RNAs that regulate gene expression by base-pairing with target mRNAs and reducing their stability or translational efficiency. Mammalian miRNAs function in association with four closely related Argonaute proteins, AGO1-4. All four proteins contain the PAZ and the MID domains interacting with the miRNA 3' and 5' termini, respectively, as well as the PIWI domain comprising an mRNA 'slicing' activity in the case of AGO2 but not AGO1, AGO3 and AGO4. However, the slicing mode of the miRNA-programmed AGO2 is rarely realized in vivo and the four Argonautes are thought to play largely overlapping roles in the mammalian miRNA pathway. Here, we show that the average length of many miRNAs is diminished during nervous system development as a result of progressive shortening of the miRNA 3' ends. We link this modification with an increase in the fractional abundance of Ago2 in the adult brain and identify a specific structural motif within the PAZ domain that enables efficient trimming of miRNAs associated with this but not the other three Argonautes. Taken together, our data suggest that mammalian Argonautes may define the length and possibly biological activity of mature mammalian miRNAs in a developmentally controlled manner.
Figures
References
-
- Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs. Annu. Rev. Biochem. 2010;79:351–379. - PubMed
-
- Kim VN, Han J, Siomi MC. Biogenesis of small RNAs in animals. Nat. Rev. Mol. Cell. Biol. 2009;10:126–139. - PubMed
-
- Ivey KN, Srivastava D. MicroRNAs as regulators of differentiation and cell fate decisions. Cell Stem Cell. 2010;7:36–41. - PubMed
-
- Fineberg SK, Kosik KS, Davidson BL. MicroRNAs potentiate neural development. Neuron. 2009;64:303–309. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
