

Genetic evidence links the ASTRA protein chaperone component Tti2 to the SAGA transcription factor Tra1
- PMID: 22505622
- PMCID: PMC3389973
- DOI: 10.1534/genetics.112.140459
Genetic evidence links the ASTRA protein chaperone component Tti2 to the SAGA transcription factor Tra1
Abstract
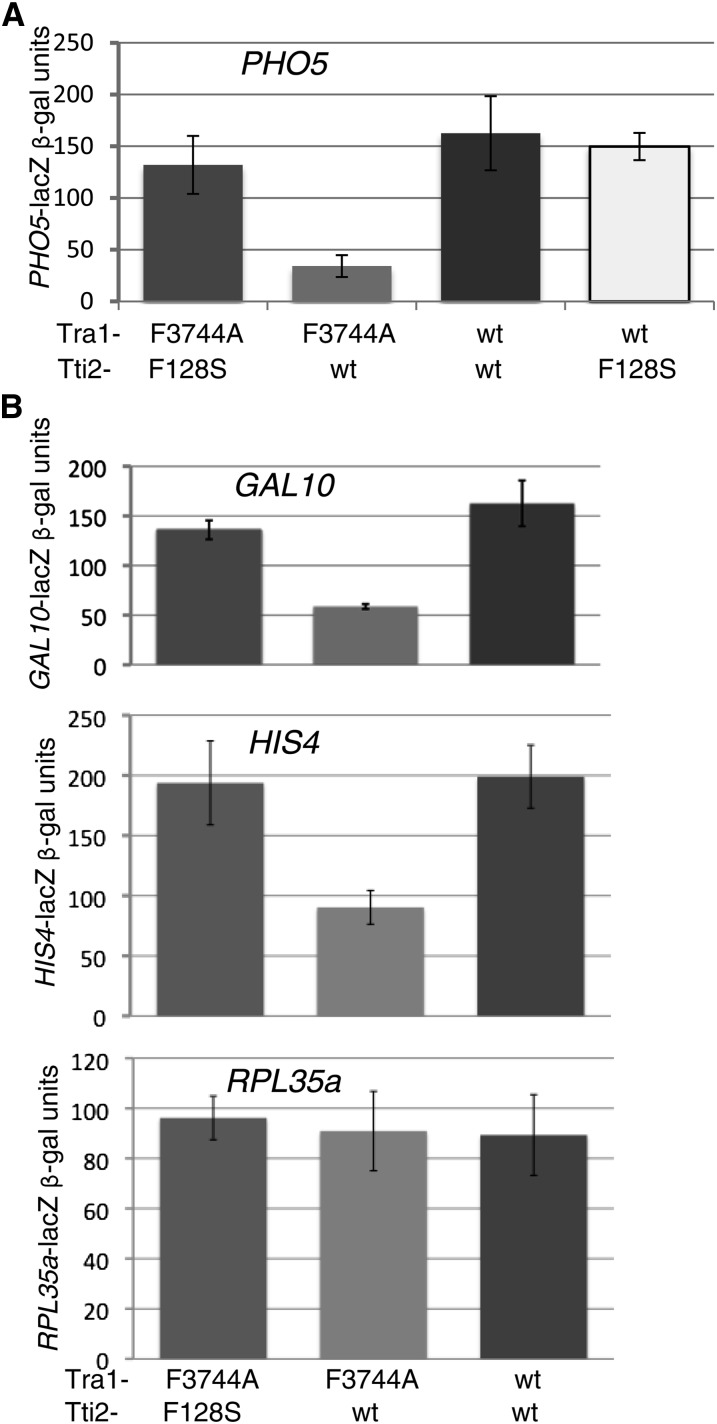
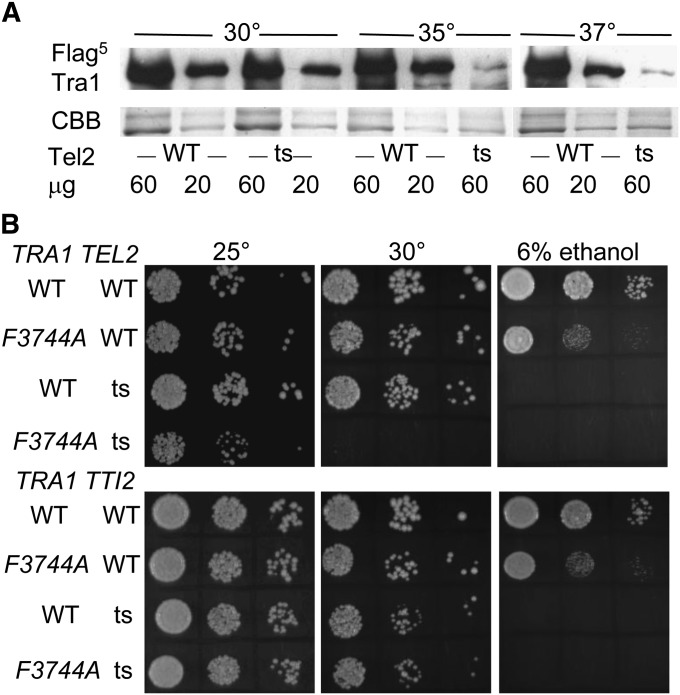
Tra1 is a 3744-residue component of the Saccharomyces cerevisiae SAGA, NuA4, and ASTRA complexes. Tra1 contains essential C-terminal PI3K and FATC domains, but unlike other PIKK (phosphoinositide three-kinase-related kinase) family members, lacks kinase activity. To analyze functions of the FATC domain, we selected for suppressors of tra1-F3744A, an allele that results in slow growth under numerous conditions of stress. Two alleles of TTI2, tti2-F328S and tti2-I336F, acted in a partially dominant fashion to suppress the growth-related phenotypes associated with tra1-F3744A as well as its resulting defects in transcription. tti2-F328S suppressed an additional FATC domain mutation (tra1-L3733A), but not a mutation in the PI3K domain or deletions of SAGA or NuA4 components. We find eGFP-tagged Tti2 distributed throughout the cell. Tti2 is a component of the ASTRA complex, and in mammalian cells associates with molecular chaperones in complex with Tti1 and Tel2. Consistent with this finding, Tra1 levels are reduced in a strain with a temperature-sensitive allele of tel2. Further agreeing with a possible role for Tti2 in the folding or stabilization of Tra1, tra1-F3744A was mislocalized to the cytoplasm, particularly under conditions of stress. Since an intragenic mutation of tra1-R3590I also suppressed F3744A, we propose that Tti2 is required for the folding/stability of the C-terminal FATC and PI3K domains of Tra1 into their functionally active form.
Figures










Similar articles
-
The Pseudokinase Domain of Saccharomyces cerevisiae Tra1 Is Required for Nuclear Localization and Incorporation into the SAGA and NuA4 Complexes.G3 (Bethesda). 2018 May 31;8(6):1943-1957. doi: 10.1534/g3.118.200288. G3 (Bethesda). 2018. PMID: 29626083 Free PMC article.
-
Mutational analysis of the C-terminal FATC domain of Saccharomyces cerevisiae Tra1.Curr Genet. 2010 Oct;56(5):447-65. doi: 10.1007/s00294-010-0313-3. Epub 2010 Jul 16. Curr Genet. 2010. PMID: 20635087 Free PMC article.
-
Loss of nonsense mediated decay suppresses mutations in Saccharomyces cerevisiae TRA1.BMC Genet. 2012 Mar 22;13:19. doi: 10.1186/1471-2156-13-19. BMC Genet. 2012. PMID: 22439631 Free PMC article.
-
Share and share alike: the role of Tra1 from the SAGA and NuA4 coactivator complexes.Transcription. 2019 Feb;10(1):37-43. doi: 10.1080/21541264.2018.1530936. Epub 2018 Oct 30. Transcription. 2019. PMID: 30375921 Free PMC article. Review.
-
Branching the Tel2 pathway for exact fit on phosphatidylinositol 3-kinase-related kinases.Curr Genet. 2018 Oct;64(5):965-970. doi: 10.1007/s00294-018-0817-9. Epub 2018 Feb 22. Curr Genet. 2018. PMID: 29470645 Free PMC article. Review.
Cited by
-
The Pseudokinase Domain of Saccharomyces cerevisiae Tra1 Is Required for Nuclear Localization and Incorporation into the SAGA and NuA4 Complexes.G3 (Bethesda). 2018 May 31;8(6):1943-1957. doi: 10.1534/g3.118.200288. G3 (Bethesda). 2018. PMID: 29626083 Free PMC article.
-
TTT (Tel2-Tti1-Tti2) Complex, the Co-Chaperone of PIKKs and a Potential Target for Cancer Chemotherapy.Int J Mol Sci. 2023 May 5;24(9):8268. doi: 10.3390/ijms24098268. Int J Mol Sci. 2023. PMID: 37175973 Free PMC article. Review.
-
Essentiality of Sis1, a J-domain protein Hsp70 cochaperone, can be overcome by Tti1, a specialized PIKK chaperone.Mol Biol Cell. 2022 Mar 1;33(3):br3. doi: 10.1091/mbc.E21-10-0493. Epub 2021 Dec 22. Mol Biol Cell. 2022. PMID: 34935410 Free PMC article.
-
The TRRAP transcription cofactor represses interferon-stimulated genes in colorectal cancer cells.Elife. 2022 Mar 4;11:e69705. doi: 10.7554/eLife.69705. Elife. 2022. PMID: 35244540 Free PMC article.
-
The phosphatidylinositol 3,5-bisphosphate (PI(3,5)P2)-dependent Tup1 conversion (PIPTC) regulates metabolic reprogramming from glycolysis to gluconeogenesis.J Biol Chem. 2013 Jul 12;288(28):20633-45. doi: 10.1074/jbc.M113.452813. Epub 2013 Jun 3. J Biol Chem. 2013. PMID: 23733183 Free PMC article.
References
-
- Abraham R. T., 2004. PI 3-kinase related kinases: ’big’ players in stress-induced signaling pathways. DNA Repair (Amst.) 3: 883–887 - PubMed
-
- Ausubel F. M., Brent R., Kingston R. E., Moore D. D., Seidman J. G., et al. , 1988. Current Protocols in Molecular Biology. Greene/Wiley-Interscience, New York, NY
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
