

Methods for quantification of in vivo changes in protein ubiquitination following proteasome and deubiquitinase inhibition
- PMID: 22505724
- PMCID: PMC3418844
- DOI: 10.1074/mcp.M111.016857
Methods for quantification of in vivo changes in protein ubiquitination following proteasome and deubiquitinase inhibition
Abstract
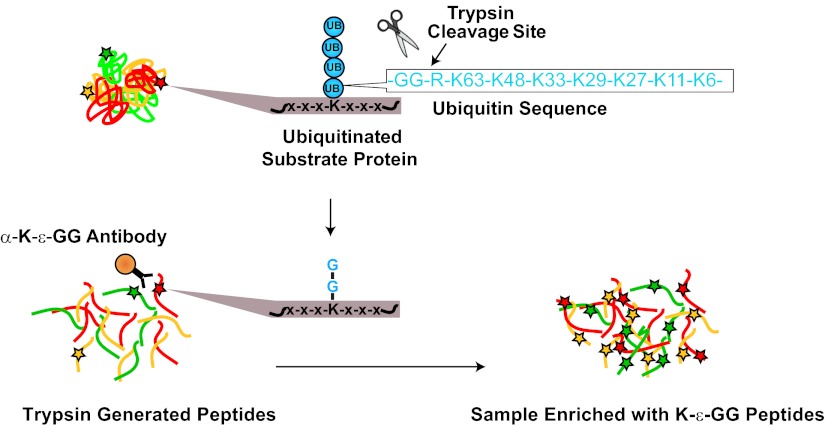
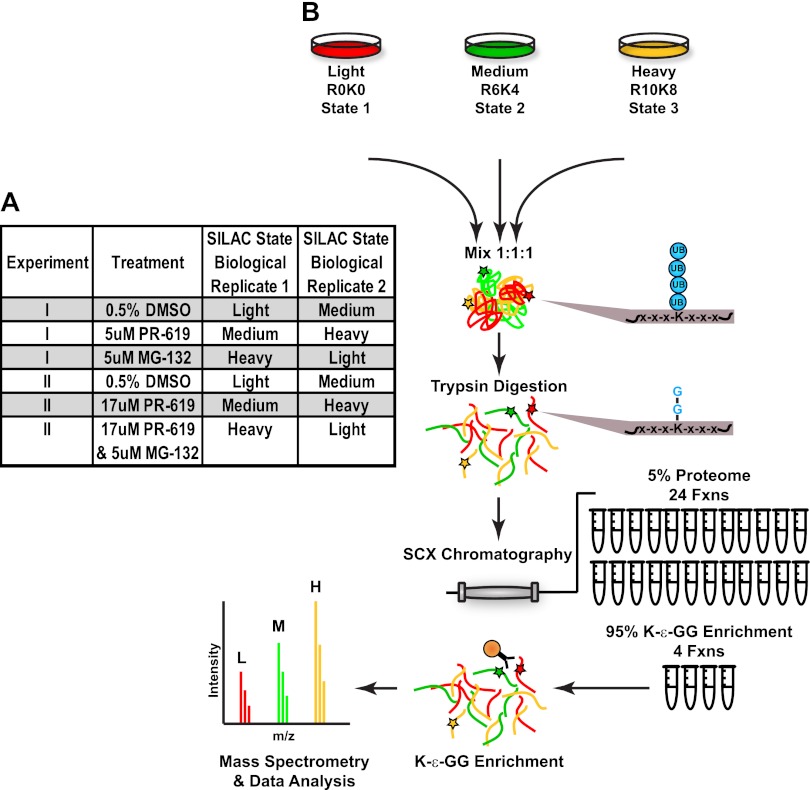
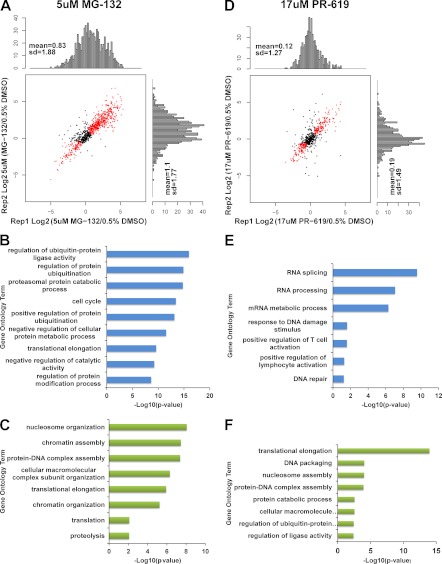
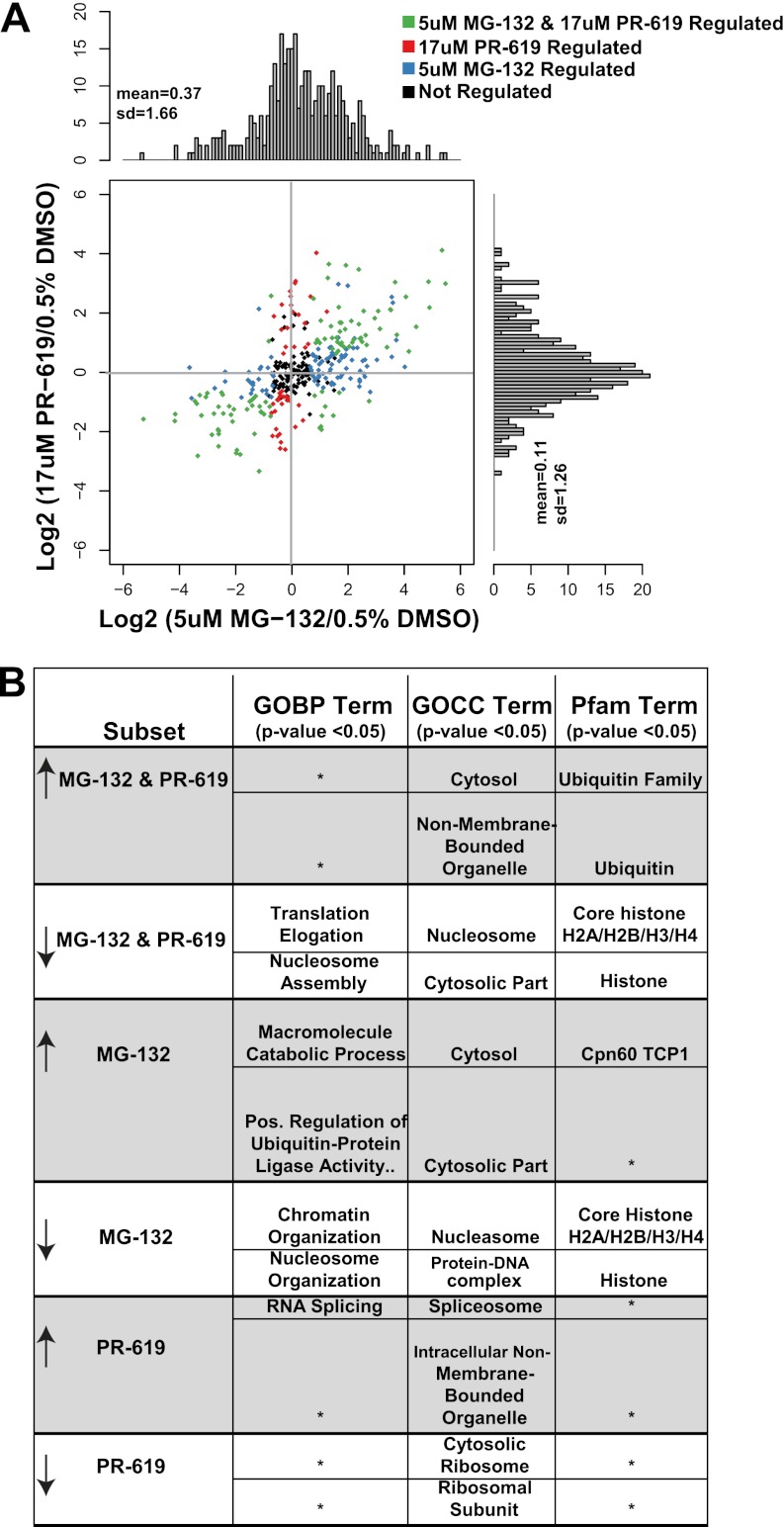
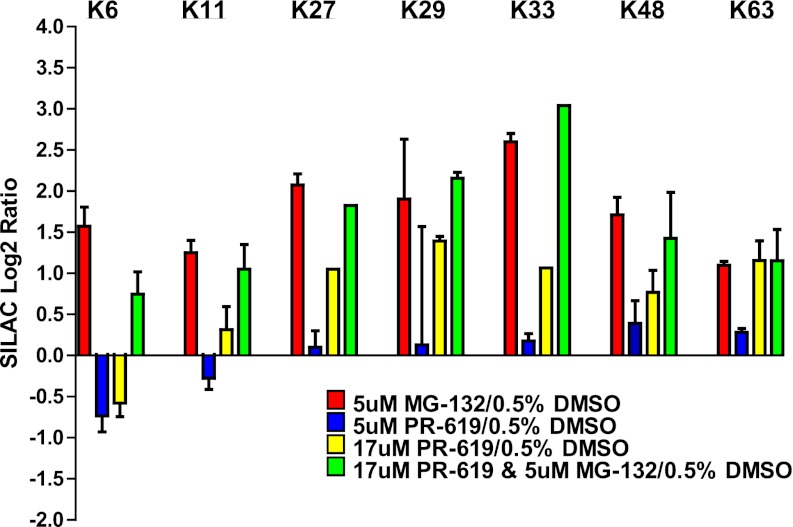
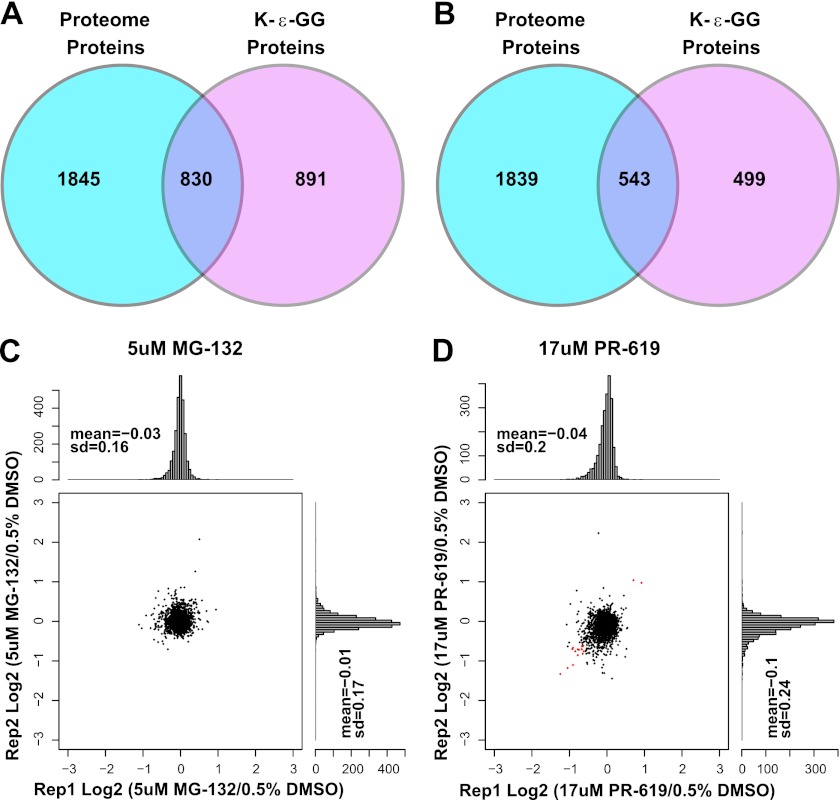
Ubiquitination plays a key role in protein degradation and signal transduction. Ubiquitin is a small protein modifier that is adducted to lysine residues by the combined function of E1, E2, and E3 enzymes and is removed by deubiquitinating enzymes. Characterization of ubiquitination sites is important for understanding the role of this modification in cellular processes and disease. However, until recently, large-scale characterization of endogenous ubiquitination sites has been hampered by the lack of efficient enrichment techniques. The introduction of antibodies that specifically recognize peptides with lysine residues that harbor a di-glycine remnant (K-ε-GG) following tryptic digestion has dramatically improved the ability to enrich and identify ubiquitination sites from cellular lysates. We used this enrichment technique to study the effects of proteasome inhibition by MG-132 and deubiquitinase inhibition by PR-619 on ubiquitination sites in human Jurkat cells by quantitative high performance mass spectrometry. Minimal fractionation of digested lysates prior to immunoaffinity enrichment increased the yield of K-ε-GG peptides three- to fourfold resulting in detection of up to ~3300 distinct K-GG peptides in SILAC triple encoded experiments starting from 5 mg of protein per label state. In total, we identify 5533 distinct K-ε-GG peptides of which 4907 were quantified in this study, demonstrating that the strategy presented is a practical approach to perturbational studies in cell systems. We found that proteasome inhibition by MG-132 and deubiquitinase inhibition by PR-619 induces significant changes to the ubiquitin landscape, but that not all ubiquitination sites regulated by MG-132 and PR-619 are likely substrates for the ubiquitin-proteasome system. Additionally, we find that the proteasome and deubiquitinase inhibitors studied induced only minor changes in protein expression levels regardless of the extent of regulation induced at the ubiquitin site level. We attribute this finding to the low stoichiometry of the majority ubiquitination sites identified in this study.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
