

A novel 3-hydroxysteroid dehydrogenase that regulates reproductive development and longevity
- PMID: 22505847
- PMCID: PMC3323522
- DOI: 10.1371/journal.pbio.1001305
A novel 3-hydroxysteroid dehydrogenase that regulates reproductive development and longevity
Abstract
Endogenous small molecule metabolites that regulate animal longevity are emerging as a novel means to influence health and life span. In C. elegans, bile acid-like steroids called the dafachronic acids (DAs) regulate developmental timing and longevity through the conserved nuclear hormone receptor DAF-12, a homolog of mammalian sterol-regulated receptors LXR and FXR. Using metabolic genetics, mass spectrometry, and biochemical approaches, we identify new activities in DA biosynthesis and characterize an evolutionarily conserved short chain dehydrogenase, DHS-16, as a novel 3-hydroxysteroid dehydrogenase. Through regulation of DA production, DHS-16 controls DAF-12 activity governing longevity in response to signals from the gonad. Our elucidation of C. elegans bile acid biosynthetic pathways reveals the possibility of novel ligands as well as striking biochemical conservation to other animals, which could illuminate new targets for manipulating longevity in metazoans.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
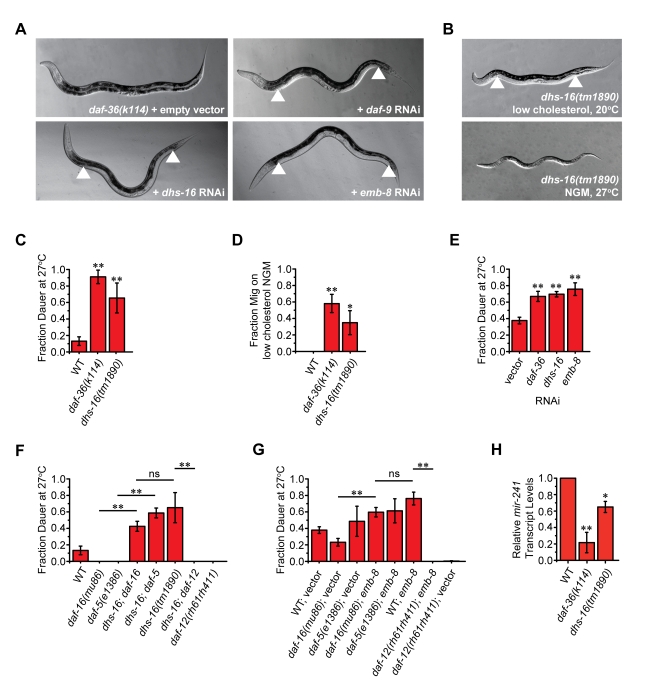
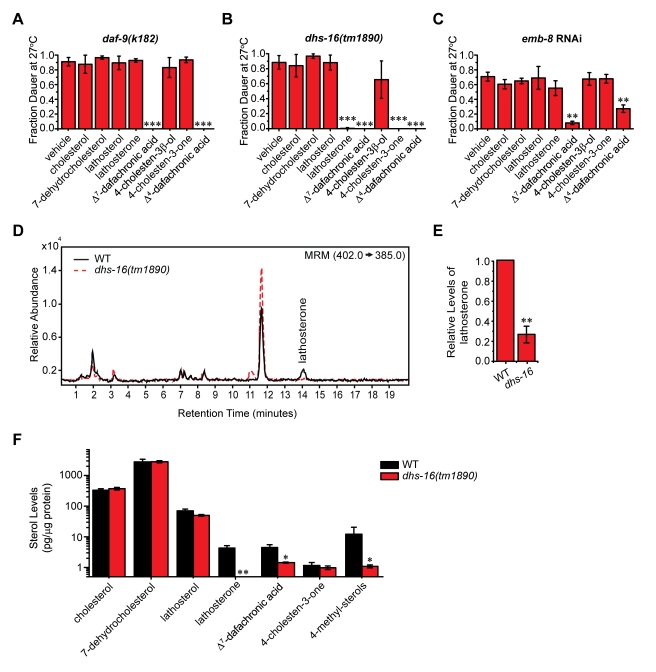
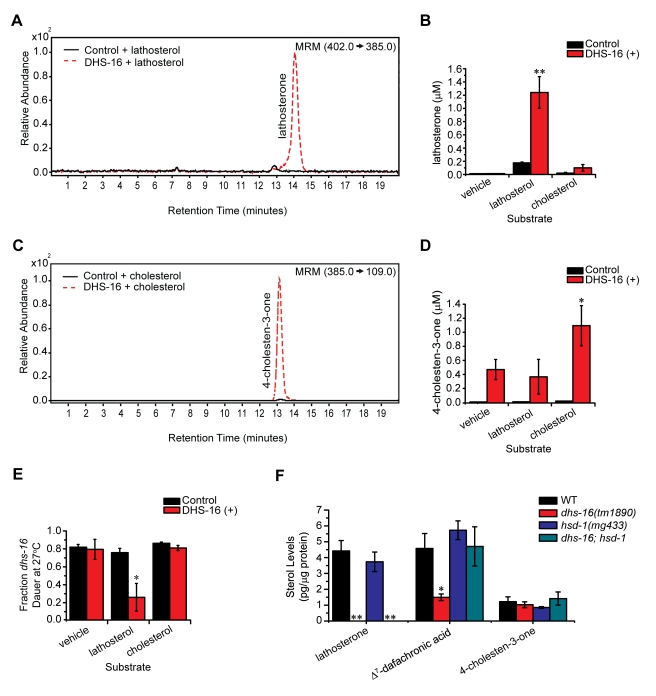
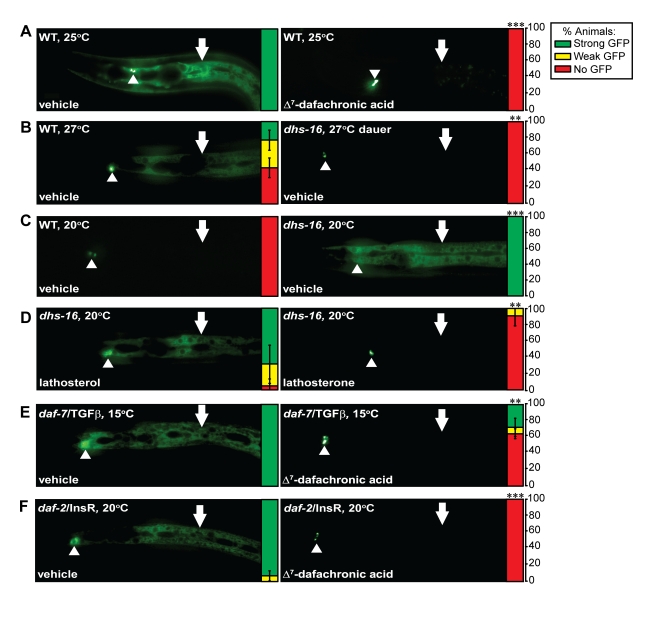
Similar articles
-
Influence of steroid hormone signaling on life span control by Caenorhabditis elegans insulin-like signaling.G3 (Bethesda). 2013 May 20;3(5):841-50. doi: 10.1534/g3.112.005116. G3 (Bethesda). 2013. PMID: 23550118 Free PMC article.
-
The Rieske oxygenase DAF-36 functions as a cholesterol 7-desaturase in steroidogenic pathways governing longevity.Aging Cell. 2011 Oct;10(5):879-84. doi: 10.1111/j.1474-9726.2011.00733.x. Epub 2011 Aug 7. Aging Cell. 2011. PMID: 21749634 Free PMC article.
-
The NHR-8 nuclear receptor regulates cholesterol and bile acid homeostasis in C. elegans.Cell Metab. 2013 Aug 6;18(2):212-24. doi: 10.1016/j.cmet.2013.07.007. Cell Metab. 2013. PMID: 23931753 Free PMC article.
-
Indicted: worms caught using steroids.Cell. 2006 Mar 24;124(6):1137-40. doi: 10.1016/j.cell.2006.03.001. Cell. 2006. PMID: 16564008 Review.
-
DAF-16: FOXO in the Context of C. elegans.Curr Top Dev Biol. 2018;127:1-21. doi: 10.1016/bs.ctdb.2017.11.007. Epub 2018 Feb 2. Curr Top Dev Biol. 2018. PMID: 29433733 Review.
Cited by
-
Activation of Nuclear Receptor CAR: A Pathway to Delay Aging through Enhanced Capacity for Xenobiotic Resistance.Adv Sci (Weinh). 2025 Mar;12(12):e2416823. doi: 10.1002/advs.202416823. Epub 2025 Jan 31. Adv Sci (Weinh). 2025. PMID: 39887667 Free PMC article.
-
No Significant Increase in the Δ4- and Δ7-Dafachronic Acid Concentration in the Long-Lived glp-1 Mutant, nor in the Mutants Defective in Dauer Formation.G3 (Bethesda). 2015 May 12;5(7):1473-9. doi: 10.1534/g3.115.018812. G3 (Bethesda). 2015. PMID: 25971936 Free PMC article.
-
Influence of steroid hormone signaling on life span control by Caenorhabditis elegans insulin-like signaling.G3 (Bethesda). 2013 May 20;3(5):841-50. doi: 10.1534/g3.112.005116. G3 (Bethesda). 2013. PMID: 23550118 Free PMC article.
-
A nematode sterol C4α-methyltransferase catalyzes a new methylation reaction responsible for sterol diversity.J Lipid Res. 2020 Feb;61(2):192-204. doi: 10.1194/jlr.RA119000317. Epub 2019 Sep 23. J Lipid Res. 2020. PMID: 31548366 Free PMC article.
-
A photocleavable masked nuclear-receptor ligand enables temporal control of C. elegans development.Angew Chem Int Ed Engl. 2014 Feb 17;53(8):2110-3. doi: 10.1002/anie.201307465. Epub 2014 Jan 22. Angew Chem Int Ed Engl. 2014. PMID: 24453122 Free PMC article.
References
-
- Kenyon C. J. The genetics of ageing. Nature. 2010;464:504–512. - PubMed
-
- Eisenberg T, Knauer H, Schauer A, Buttner S, Ruckenstuhl C, et al. Induction of autophagy by spermidine promotes longevity. Nat Cell Biol. 2009;11:1305–1314. - PubMed
-
- Honda Y, Tanaka M, Honda S. Trehalose extends longevity in the nematode Caenorhabditis elegans. Aging Cell. 2010;9:558–569. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
