

doi: 10.1155/2012/941292.
Epub 2012 Mar 8.
Actin dynamics associated with focal adhesions
Affiliations
- PMID: 22505938
- PMCID: PMC3312244
- DOI: 10.1155/2012/941292
Item in Clipboard
Actin dynamics associated with focal adhesions
Int J Cell Biol.
2012.
Abstract
Cell-matrix adhesion plays a major role during cell migration. Proteins from adhesion structures connect the extracellular matrix to the actin cytoskeleton, allowing the growing actin network to push the plasma membrane and the contractile cables (stress fibers) to pull the cell body. Force transmission to the extracellular matrix depends on several parameters including the regulation of actin dynamics in adhesion structures, the contractility of stress fibers, and the mechanosensitive response of adhesion structures. Here we highlight recent findings on the molecular mechanisms by which actin assembly is regulated in adhesion structures and the molecular basis of the mechanosensitivity of focal adhesions.
Figures
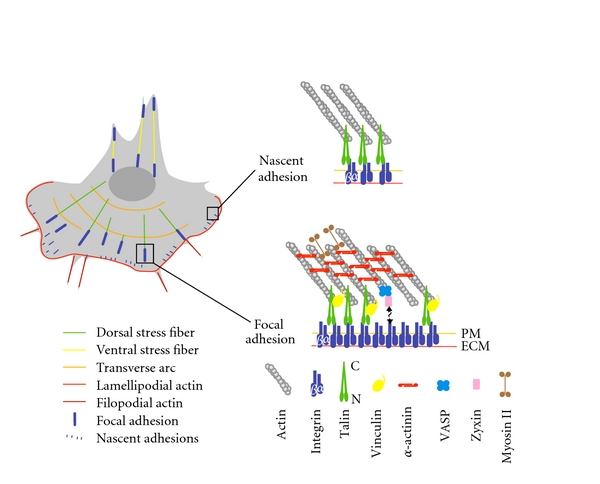
Actin networks in cell migration and organization of nascent adhesions and focal adhesions. Left, scheme of a migrating cell displaying characteristic actin structures: lamellipodial and filopodial actin networks and the three classes of stress fibers (transverse arcs, dorsal stress fibers, ventral stress fibers). Right, actin-binding proteins in focal complexes and focal adhesions. PM, plasma membrane; ECM, extracellular matrix.
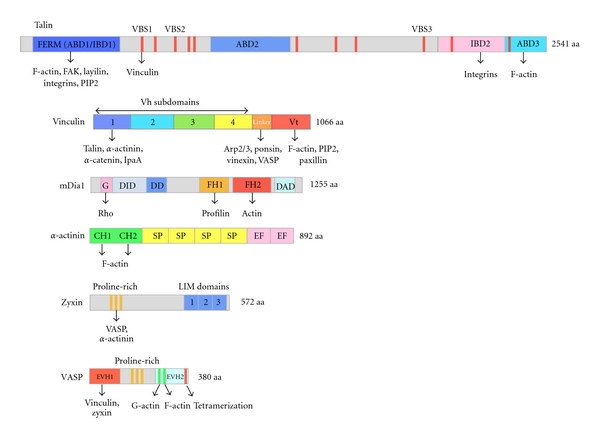
Domain organization of proteins involved in the regulation of actin assembly in focal adhesions. ABD, actin-binding domain; FERM, four-point-one, ezrin, radixin, moesin; IBD, integrin binding domain; VBS, vinculin-binding site (11 VBSs are indicated as red vertical bars); GBD, G-binding domain; DID, diaphanous inhibitory domain; DD, dimerization domain; FH1, formin homology domain 1; FH2, formin homology domain 2; DAD, diaphanous autoregulatory domain; CH, calponin homology; SP, spectrin repeat; EF, EF hand motif; EVH1, Ena/VASP homology domain 1; EVH2, Ena/VASP homology domain 2.
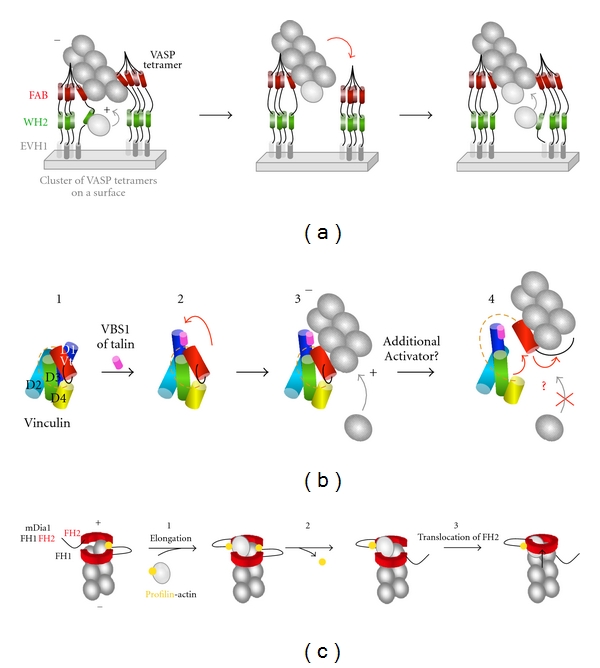
Regulation of actin assembly by VASP, vinculin, and formins. (a) VASP-mediated processive elongation of actin filaments. Clusters of VASP tetramers allow the WH2 domains of several VASP molecules to deliver actin monomers to actin filament barbed ends in a processive manner. This activity is accompanied by an acceleration of barbed end elongation. Between delivery events, VASP remains associated to the side of the filament via its FAB domain, adapted from [42, 43]. (+) and (−) indicate the barbed end and the pointed end of the actin filament. (b) Regulation of actin filament side binding and barbed-end capping by vinculin. In this scheme, subdomains of vinculin are represented as cylinders. The proline-rich region that links Vh and Vt is represented as an orange dotted line. (1) Vinculin is autoinhibited by an intramolecular interaction between Vh (D1 to D4) and Vt. The F-actin binding site located in Vt is masked by D1. (2) The binding of the VBS1 domain of talin disrupts the D1-Vt interaction (red arrow). Vt is unmasked and binds to the side of an actin filament. Barbed end elongation is still possible. (4) The disruption of additional unidentified contacts unmasks the C-terminal arm (black line) of Vt which caps the barbed end of the filament, adapted from [41]. (c) Formin-mediated processive elongation of actin filaments. In this scheme, only the FH1 (black line) and FH2 (red) domains are represented. (1) Addition of a profilin-actin subunit to the formin-bound barbed end. Each FH2 protomer of the formin dimer makes two contacts with the terminal actin subunits at a barbed end. (2) Profilin dissociates. (3) The translocation of one FH2 protomer releases one of the two actin-formin bonds to allow the addition of next profilin-actin subunit, adapted from [3].
Similar articles
-
Actin machinery and mechanosensitivity in invadopodia, podosomes and focal adhesions.J Cell Sci. 2009 Sep 1;122(Pt 17):3037-49. doi: 10.1242/jcs.052704. J Cell Sci. 2009. PMID: 19692590 Free PMC article. Review.
-
Stressing the limits of focal adhesion mechanosensitivity.Curr Opin Cell Biol. 2014 Oct;30:68-73. doi: 10.1016/j.ceb.2014.06.003. Epub 2014 Jul 5. Curr Opin Cell Biol. 2014. PMID: 24998185 Free PMC article. Review.
-
Generation of contractile actomyosin bundles depends on mechanosensitive actin filament assembly and disassembly.Elife. 2015 Dec 10;4:e06126. doi: 10.7554/eLife.06126. Elife. 2015. PMID: 26652273 Free PMC article.
-
Force-induced adsorption and anisotropic growth of focal adhesions.Biophys J. 2006 May 15;90(10):3469-84. doi: 10.1529/biophysj.105.074377. Epub 2006 Mar 2. Biophys J. 2006. PMID: 16513789 Free PMC article.
-
Early molecular events in the assembly of the focal adhesion-stress fiber complex during fibroblast spreading.Cell Motil Cytoskeleton. 2004 Jul;58(3):143-59. doi: 10.1002/cm.20005. Cell Motil Cytoskeleton. 2004. PMID: 15146534
Cited by
-
RhoA activation promotes glucose uptake to elevate proliferation in MAPK inhibitor resistant melanoma cells.bioRxiv [Preprint]. 2024 Jan 11:2024.01.09.574940. doi: 10.1101/2024.01.09.574940. bioRxiv. 2024. PMID: 38260449 Free PMC article. Preprint.
-
Dynamic compartmentalization of purine nucleotide metabolic enzymes at leading edge in highly motile renal cell carcinoma.Biochem Biophys Res Commun. 2019 Aug 13;516(1):50-56. doi: 10.1016/j.bbrc.2019.05.190. Epub 2019 Jun 10. Biochem Biophys Res Commun. 2019. PMID: 31196624 Free PMC article.
-
Promotion of Melanoma Cell Proliferation by Cyclic Straining through Regulatory Morphogenesis.Int J Mol Sci. 2022 Oct 6;23(19):11884. doi: 10.3390/ijms231911884. Int J Mol Sci. 2022. PMID: 36233186 Free PMC article.
-
A Focal Adhesion Filament Cross-correlation Kit for fast, automated segmentation and correlation of focal adhesions and actin stress fibers in cells.PLoS One. 2021 Sep 10;16(9):e0250749. doi: 10.1371/journal.pone.0250749. eCollection 2021. PLoS One. 2021. PMID: 34506490 Free PMC article.
-
The Arp2/3 complex is crucial for colonisation of the mouse skin by melanoblasts.Development. 2020 Nov 15;147(22):dev194555. doi: 10.1242/dev.194555. Development. 2020. PMID: 33028610 Free PMC article.
References
-
- Wang W, Goswami S, Sahai E, Wyckoff JB, Segall JE, Condeelis JS. Tumor cells caught in the act of invading: their strategy for enhanced cell motility. Trends in Cell Biology. 2005;15(3):138–145. - PubMed
-
- Ridley AJ, Schwartz MA, Burridge K, et al. Cell migration: integrating signals from front to back. Science. 2003;302(5651):1704–1709. - PubMed
-
- Le Clainche C, Carlier MF. Regulation of actin assembly associated with protrusion and adhesion in cell migration. Physiological Reviews. 2008;88(2):489–513. - PubMed
-
- Critchley DR. Biochemical and structural properties of the integrin-associated cytoskeletal protein talin. Annual Review of Biophysics. 2009;38(1):235–254. - PubMed
-
- Zaidel-Bar R, Ballestrem C, Kam Z, Geiger B. Early molecular events in the assembly of matrix adhesions at the leading edge of migrating cells. Journal of Cell Science. 2003;116, part 22:4605–4613. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
