

MicroRNAs overexpressed in growth-restricted rat skeletal muscles regulate the glucose transport in cell culture targeting central TGF-β factor SMAD4
- PMID: 22506032
- PMCID: PMC3323545
- DOI: 10.1371/journal.pone.0034596
MicroRNAs overexpressed in growth-restricted rat skeletal muscles regulate the glucose transport in cell culture targeting central TGF-β factor SMAD4
Abstract
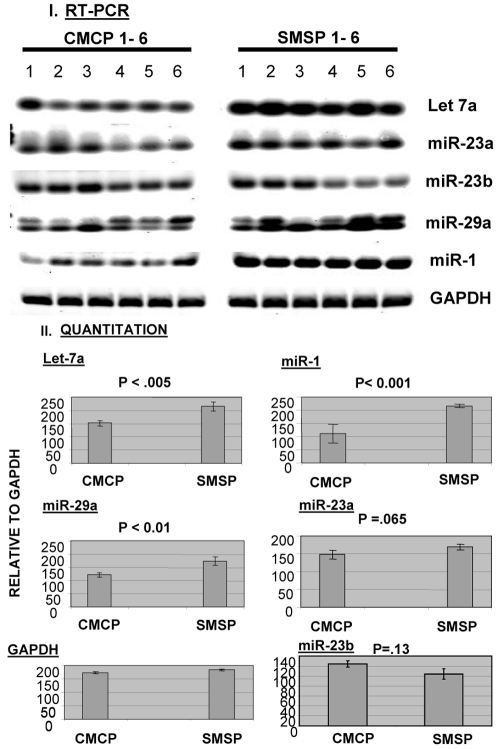
The micro-array profiling of micro-RNA has been performed in rat skeletal muscle tissues, isolated from male adult offspring of intrauterine plus postnatal growth restricted model (IPGR). Apparently, the GLUT4 mRNA expression in male sk. muscle was found to be unaltered in contrast to females. The over-expression of miR-29a and miR-23a in the experimental group of SMSP (Starved Mother Starved Pups) have been found to regulate the glucose transport activity with respect to their control counterparts CMCP (Control Mother Control Pups) as confirmed in rat L6 myoblast-myocyte cell culture system. The ex-vivo experimentation demonstrates an aberration in insulin signaling pathway in male sk. muscle that leads to the localization of the membrane-bound Glut4 protein. We have identified through a series of experiments one important protein factor SMAD4, a co-SMAD critical to the TGF-beta signaling pathway. This factor is targeted by miR-29a, as identified in an in vitro reporter-assay system in cell-culture experiment. The other micro-RNA, miR-23a, targets SMAD4 indirectly that seems to be critical in regulating insulin-dependent glucose transport activity. MicroRNA mimics, inhibitors and siRNA studies indicate the role of SMAD4 as inhibitory for glucose transport activities in normal physiological condition. The data demonstrate for the first time a critical function of microRNAs in fine-tuning the regulation of glucose transport in skeletal muscle. Chronic starved conditions (IPGR) in sk. muscle up-regulates microRNA changing the target protein expression patterns, such as SMAD4, to alter the glucose transport pathways for the survival. The innovative outcome of this paper identifies a critical pathway (TGF-beta) that may act negatively for the mammalian glucose transport machinery.
Conflict of interest statement
Figures













References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
