

Lack of evidence from studies of soluble protein fragments that Knops blood group polymorphisms in complement receptor-type 1 are driven by malaria
- PMID: 22506052
- PMCID: PMC3323580
- DOI: 10.1371/journal.pone.0034820
Lack of evidence from studies of soluble protein fragments that Knops blood group polymorphisms in complement receptor-type 1 are driven by malaria
Abstract
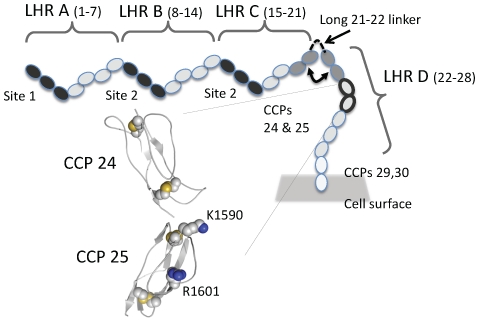
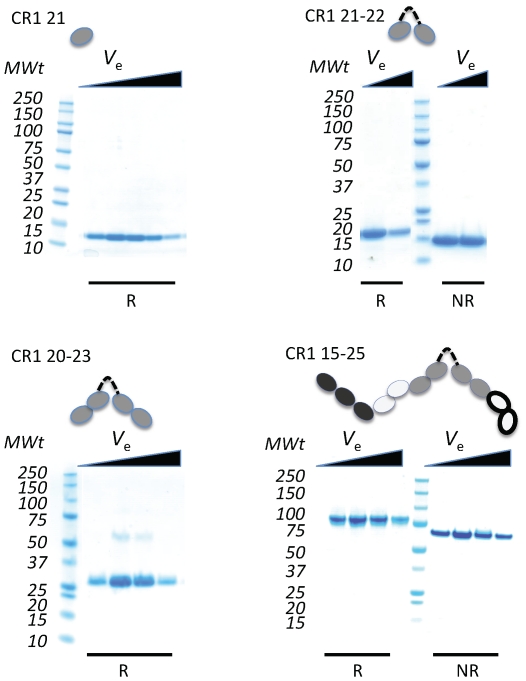
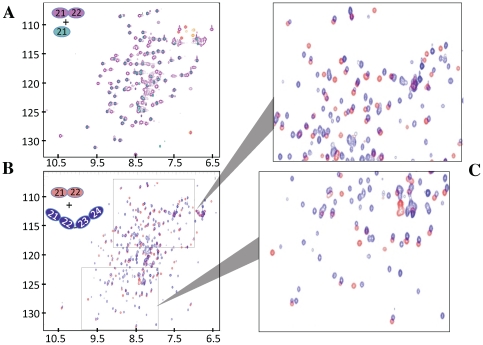
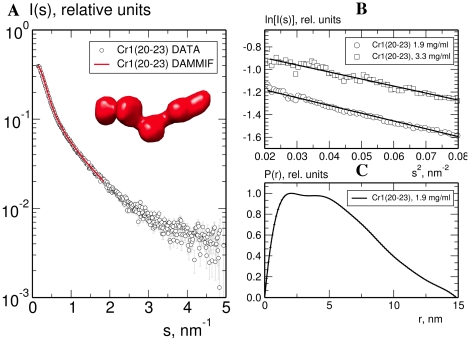
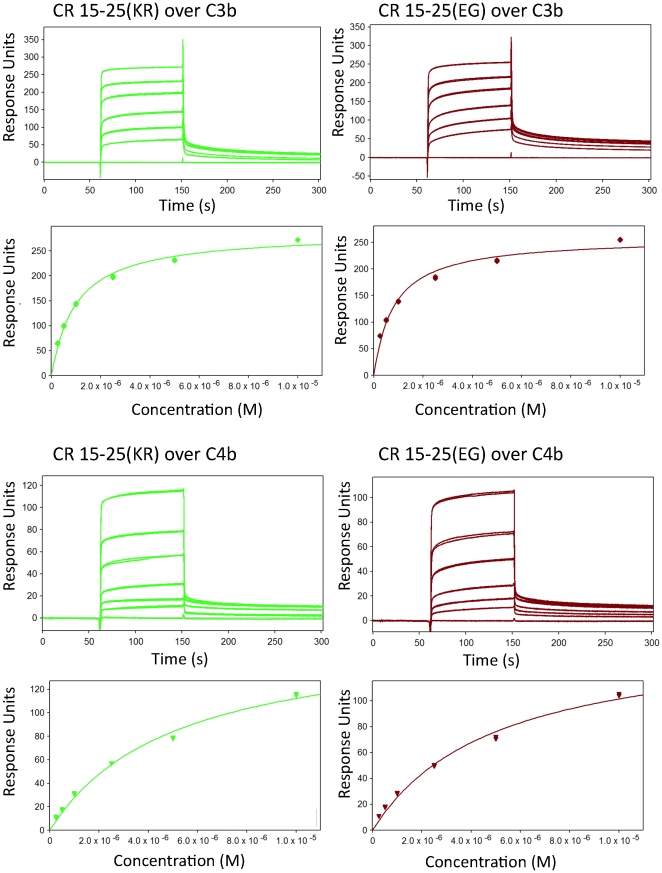
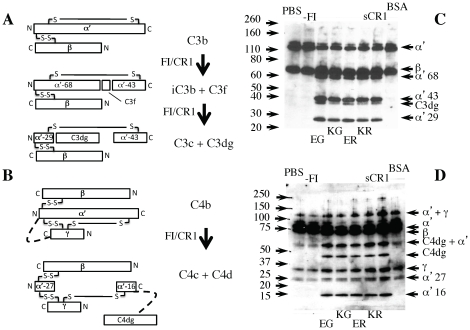
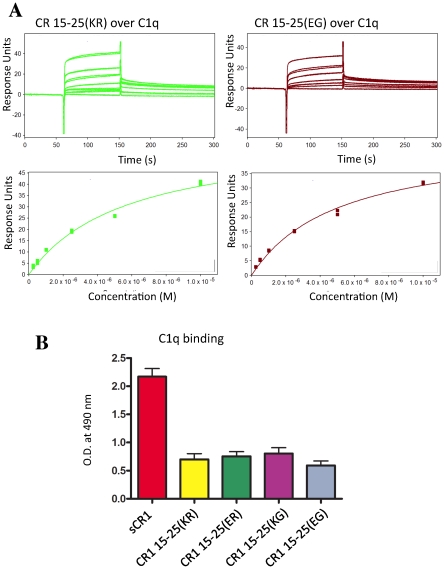
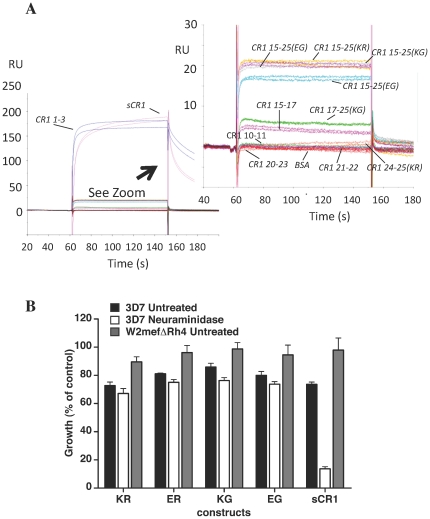
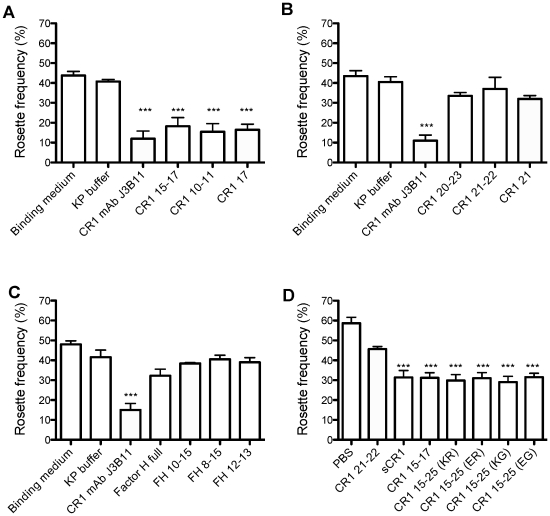
Complement receptor-type 1 (CR1, CD35) is the immune-adherence receptor, a complement regulator, and an erythroid receptor for Plasmodium falciparum during merozoite invasion and subsequent rosette formation involving parasitized and non-infected erythrocytes. The non-uniform geographical distribution of Knops blood group CR1 alleles Sl1/2 and McC(a/b) may result from selective pressures exerted by differential exposure to infectious hazards. Here, four variant short recombinant versions of CR1 were produced and analyzed, focusing on complement control protein modules (CCPs) 15-25 of its ectodomain. These eleven modules encompass a region (CCPs 15-17) key to rosetting, opsonin recognition and complement regulation, as well as the Knops blood group polymorphisms in CCPs 24-25. All four CR1 15-25 variants were monomeric and had similar axial ratios. Modules 21 and 22, despite their double-length inter-modular linker, did not lie side-by-side so as to stabilize a bent-back architecture that would facilitate cooperation between key functional modules and Knops blood group antigens. Indeed, the four CR1 15-25 variants had virtually indistinguishable affinities for immobilized complement fragments C3b (K(D) = 0.8-1.1 µM) and C4b (K(D) = 5.0-5.3 µM). They were all equally good co-factors for factor I-catalysed cleavage of C3b and C4b, and they bound equally within a narrow affinity range, to immobilized C1q. No differences between the variants were observed in assays for inhibition of erythrocyte invasion by P. falciparum or for rosette disruption. Neither differences in complement-regulatory functionality, nor interactions with P. falciparum proteins tested here, appear to have driven the non-uniform geographic distribution of these alleles.
Conflict of interest statement
Figures
Similar articles
-
Using mutagenesis and structural biology to map the binding site for the Plasmodium falciparum merozoite protein PfRh4 on the human immune adherence receptor.J Biol Chem. 2014 Jan 3;289(1):450-63. doi: 10.1074/jbc.M113.520346. Epub 2013 Nov 8. J Biol Chem. 2014. PMID: 24214979 Free PMC article.
-
CR1 Knops blood group alleles are not associated with severe malaria in the Gambia.Genes Immun. 2003 Jul;4(5):368-73. doi: 10.1038/sj.gene.6363980. Genes Immun. 2003. PMID: 12847553 Free PMC article.
-
Molecular identification of Knops blood group polymorphisms found in long homologous region D of complement receptor 1.Blood. 2001 May 1;97(9):2879-85. doi: 10.1182/blood.v97.9.2879. Blood. 2001. PMID: 11313284
-
[The structure and role of CR1 complement receptor in physiology].Pol Merkur Lekarski. 2010 Jan;28(163):79-83. Pol Merkur Lekarski. 2010. PMID: 20369732 Review. Polish.
-
A Review of the Knops Blood Group System.Clin Appl Thromb Hemost. 2024 Jan-Dec;30:10760296241309638. doi: 10.1177/10760296241309638. Clin Appl Thromb Hemost. 2024. PMID: 39706812 Free PMC article. Review.
Cited by
-
Role of complement receptor 1 (CR1; CD35) on epithelial cells: A model for understanding complement-mediated damage in the kidney.Mol Immunol. 2015 Oct;67(2 Pt B):584-95. doi: 10.1016/j.molimm.2015.07.016. Epub 2015 Aug 7. Mol Immunol. 2015. PMID: 26260209 Free PMC article.
-
No Evidence that Knops Blood Group Polymorphisms Affect Complement Receptor 1 Clustering on Erythrocytes.Sci Rep. 2017 Dec 19;7(1):17825. doi: 10.1038/s41598-017-17664-9. Sci Rep. 2017. PMID: 29259218 Free PMC article.
-
Blood Groups in Infection and Host Susceptibility.Clin Microbiol Rev. 2015 Jul;28(3):801-70. doi: 10.1128/CMR.00109-14. Clin Microbiol Rev. 2015. PMID: 26085552 Free PMC article. Review.
-
Analysis of the Putative Role of CR1 in Alzheimer's Disease: Genetic Association, Expression and Function.PLoS One. 2016 Feb 25;11(2):e0149792. doi: 10.1371/journal.pone.0149792. eCollection 2016. PLoS One. 2016. PMID: 26914463 Free PMC article.
-
C1q and Mannose-Binding Lectin Interact with CR1 in the Same Region on CCP24-25 Modules.Front Immunol. 2018 Mar 7;9:453. doi: 10.3389/fimmu.2018.00453. eCollection 2018. Front Immunol. 2018. PMID: 29563915 Free PMC article.
References
-
- Moulds JM. A review of the Knops blood group: separating fact from fallacy. Immunohematology. 2002;18:1–8. - PubMed
-
- Moulds JM. The Knops blood-group system: a review. Immunohematology. 2010;26:2–7. - PubMed
-
- Moulds JM, Zimmerman PA, Doumbo OK, Kassambara L, Sagara I, et al. Molecular identification of Knops blood group polymorphisms found in long homologous region D of complement receptor 1. Blood. 2001;97:2879–2885. - PubMed
-
- Stoute JA. Complement-regulatory proteins in severe malaria: too little or too much of a good thing? Trends Parasitol. 2005;21:218–223. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
