

GABA(A) receptors containing the α2 subunit are critical for direction-selective inhibition in the retina
- PMID: 22506070
- PMCID: PMC3323634
- DOI: 10.1371/journal.pone.0035109
GABA(A) receptors containing the α2 subunit are critical for direction-selective inhibition in the retina
Abstract
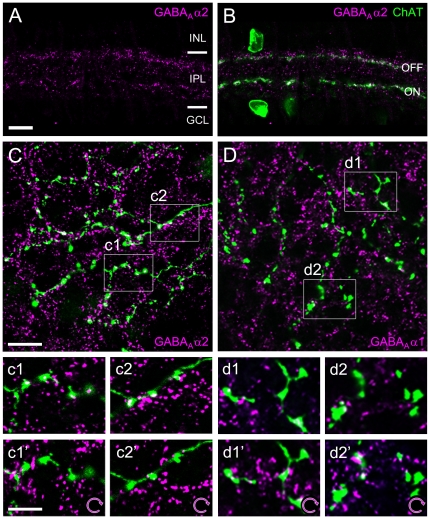
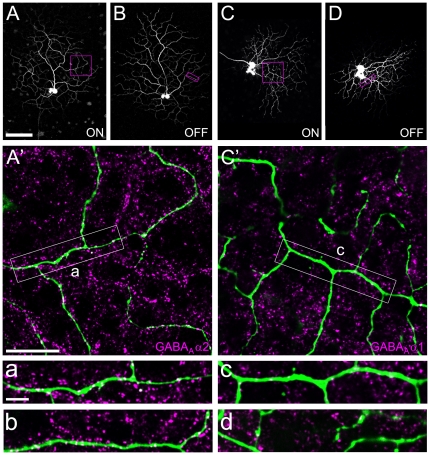
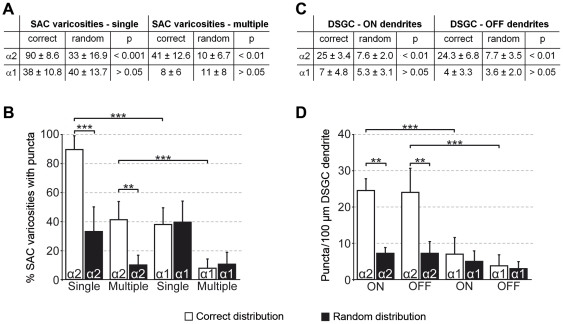
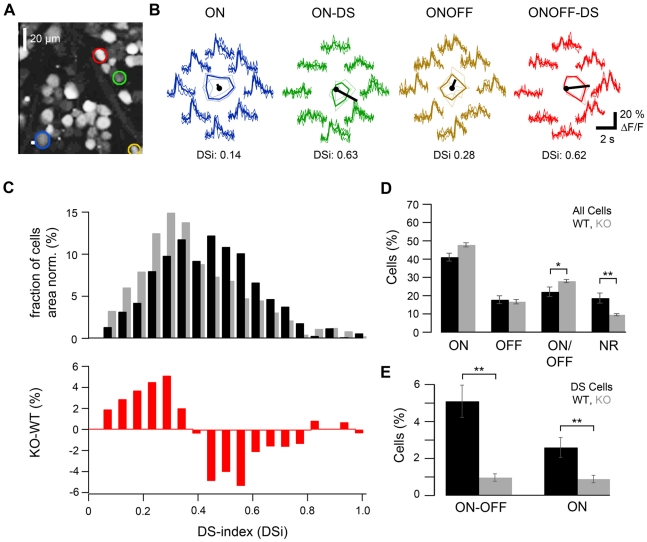
Far from being a simple sensor, the retina actively participates in processing visual signals. One of the best understood aspects of this processing is the detection of motion direction. Direction-selective (DS) retinal circuits include several subtypes of ganglion cells (GCs) and inhibitory interneurons, such as starburst amacrine cells (SACs). Recent studies demonstrated a surprising complexity in the arrangement of synapses in the DS circuit, i.e. between SACs and DS ganglion cells. Thus, to fully understand retinal DS mechanisms, detailed knowledge of all synaptic elements involved, particularly the nature and localization of neurotransmitter receptors, is needed. Since inhibition from SACs onto DSGCs is crucial for generating retinal direction selectivity, we investigate here the nature of the GABA receptors mediating this interaction. We found that in the inner plexiform layer (IPL) of mouse and rabbit retina, GABA(A) receptor subunit α2 (GABA(A)R α2) aggregated in synaptic clusters along two bands overlapping the dendritic plexuses of both ON and OFF SACs. On distal dendrites of individually labeled SACs in rabbit, GABA(A)R α2 was aligned with the majority of varicosities, the cell's output structures, and found postsynaptically on DSGC dendrites, both in the ON and OFF portion of the IPL. In GABA(A)R α2 knock-out (KO) mice, light responses of retinal GCs recorded with two-photon calcium imaging revealed a significant impairment of DS responses compared to their wild-type littermates. We observed a dramatic drop in the proportion of cells exhibiting DS phenotype in both the ON and ON-OFF populations, which strongly supports our anatomical findings that α2-containing GABA(A)Rs are critical for mediating retinal DS inhibition. Our study reveals for the first time, to the best of our knowledge, the precise functional localization of a specific receptor subunit in the retinal DS circuit.
Conflict of interest statement
Figures
References
-
- Barlow HB, Hill RM. Selective sensitivity to direction of movement in ganglion cells of the rabbit retina. Science. 1963;139:412–414. - PubMed
-
- Amthor FR, Oyster CW, Takahashi ES. Morphology of on-off direction-selective ganglion cells in the rabbit retina. Brain Res. 1984;298:187–190. - PubMed
-
- Amthor FR, Takahashi ES, Oyster CW. Morphologies of rabbit retinal ganglion cells with complex receptive fields. J Comp Neurol. 1989;280:97–121. - PubMed
-
- Oyster CW, Barlow HB. Direction-selective units in rabbit retina: distribution of preferred directions. Science. 1967;155:841–842. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
