

Augmentation of cognitive function by NS9283, a stoichiometry-dependent positive allosteric modulator of α2- and α4-containing nicotinic acetylcholine receptors
- PMID: 22506660
- PMCID: PMC3448921
- DOI: 10.1111/j.1476-5381.2012.01989.x
Augmentation of cognitive function by NS9283, a stoichiometry-dependent positive allosteric modulator of α2- and α4-containing nicotinic acetylcholine receptors
Abstract
Background and purpose: Positive allosteric modulation of α4β2 nicotinic acetylcholine (nACh) receptors could add a new dimension to the pharmacology and therapeutic approach to these receptors. The novel modulator NS9283 was therefore tested extensively.
Experimental approach: Effects of NS9283 were evaluated in vitro using fluorescence-based Ca(2+) imaging and electrophysiological voltage clamp experiments in Xenopus oocytes, mammalian cells and thalamocortical neurons. In vivo the compound was tested in models covering a range of cognitive domains in mice and rats.
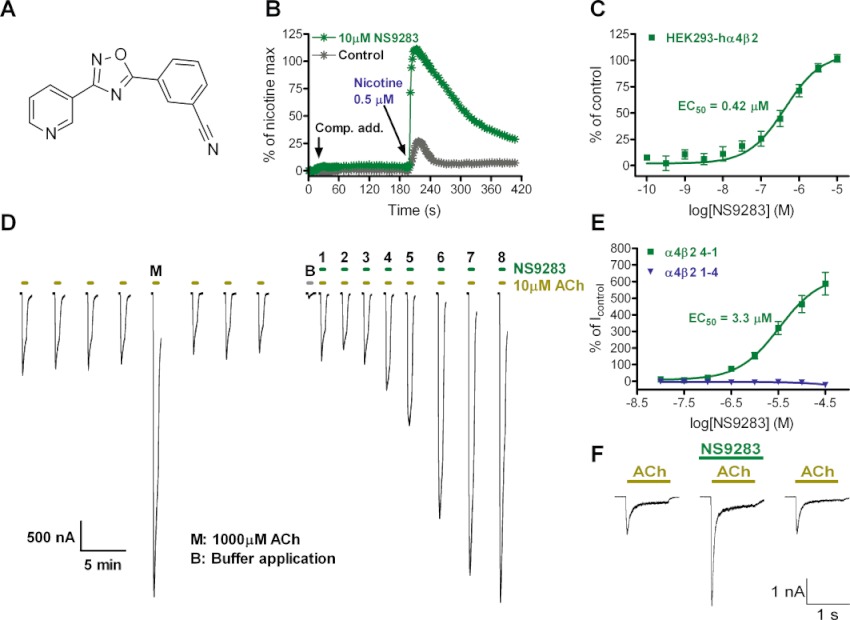
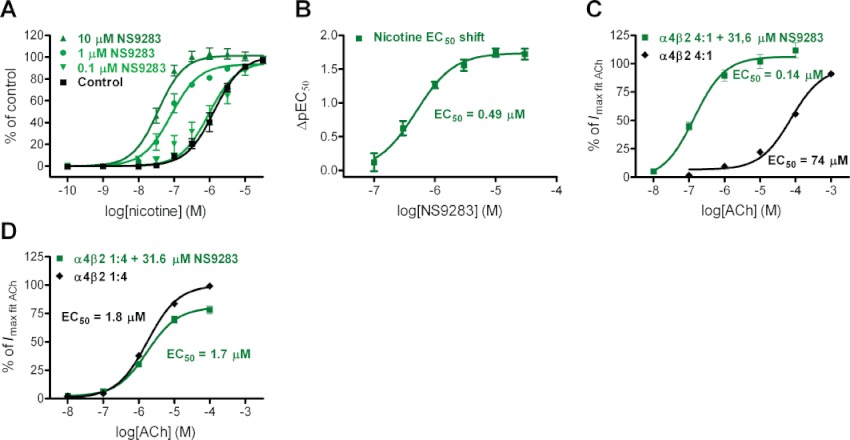
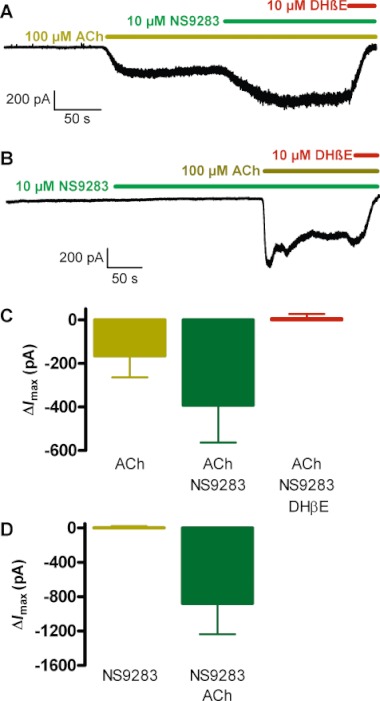
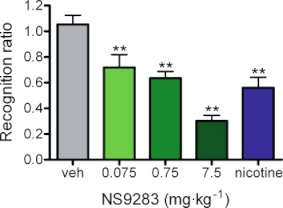
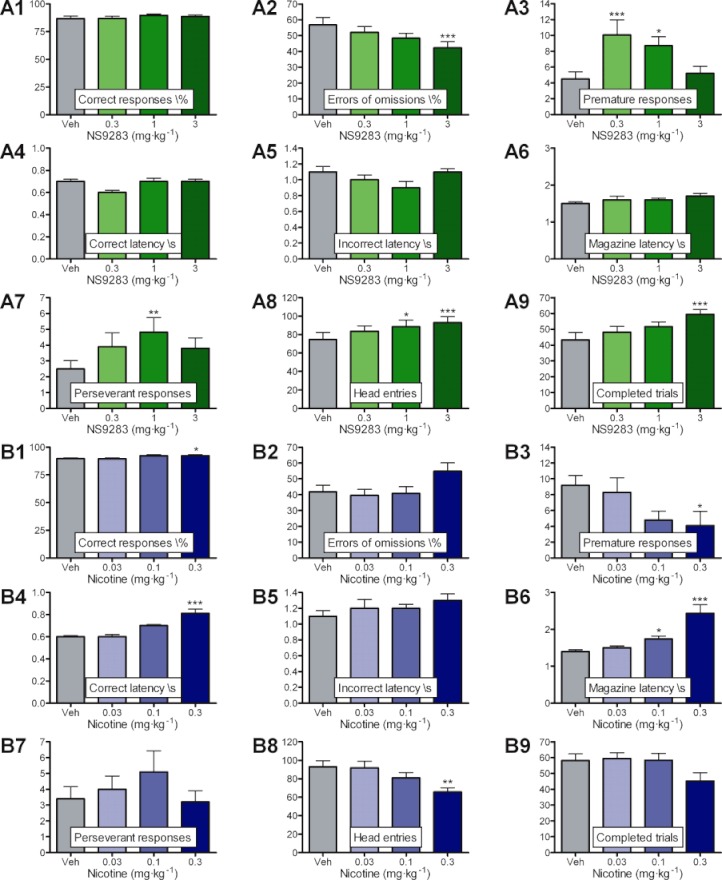
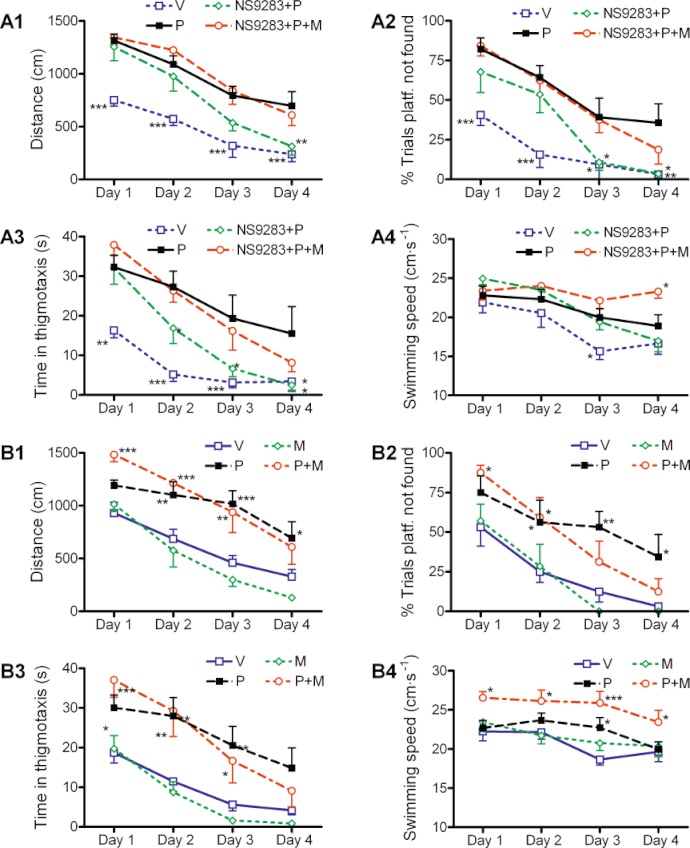
Key results: NS9283 was shown to increase agonist-evoked response amplitude of (α4)(3) (β2)(2) nACh receptors in electrophysiology paradigms. (α2)(3) (β2)(2) , (α2)(3) (β4)(2) and (α4)(3) (β4)(2) were modulated to comparable extents, but no effects were detected at α3-containing or any 2α : 3β stoichiometry nACh receptors. Native nACh receptors in thalamocortical neurons similarly displayed DHβE-sensitive currents that were receptive to modulation. NS9283 had favourable effects on sensory information processing, as shown by reversal of PCP-disrupted pre-pulse inhibition. NS9283 further improved performance in a rat model of episodic memory (social recognition), a rat model of sustained attention (five-choice serial reaction time task) and a rat model of reference memory (Morris water maze). Importantly, the effects in the Morris water maze could be fully reversed with mecamylamine, a blocker of nACh receptors.
Conclusions and implications: These results provide compelling evidence that positive allosteric modulators acting at the (α4)(3) (β2)(2) nACh receptors can augment activity across a broad range of cognitive domains, and that α4β2 nACh receptor allosteric modulation therefore constitutes a promising therapeutic approach to symptomatic treatment of cognitive impairment.
© 2012 The Authors. British Journal of Pharmacology © 2012 The British Pharmacological Society.
Figures
References
-
- Acri JB, Morse DE, Popke EJ, Grunberg NE. Nicotine increases sensory gating measured as inhibition of the acoustic startle reflex in rats. Psychopharmacology (Berl) 1994;114:369–374. - PubMed
-
- Albrecht BK, Berry V, Boezio AA, Cao L, Clarkin K, Guo W, et al. Discovery and optimization of substituted piperidines as potent, selective, CNS-penetrant alpha4beta2 nicotinic acetylcholine receptor potentiators. Bioorg Med Chem Lett. 2008;18:5209–5212. - PubMed
-
- Briggs CA, Gubbins EJ, Marks MJ, Putman CB, Thimmapaya R, Meyer MD, et al. Untranslated region-dependent exclusive expression of high-sensitivity subforms of alpha4beta2 and alpha3beta2 nicotinic acetylcholine receptors. Mol Pharmacol. 2006;70:227–240. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
