

Munc13-4 reconstitutes calcium-dependent SNARE-mediated membrane fusion
- PMID: 22508512
- PMCID: PMC3328385
- DOI: 10.1083/jcb.201109132
Munc13-4 reconstitutes calcium-dependent SNARE-mediated membrane fusion
Abstract
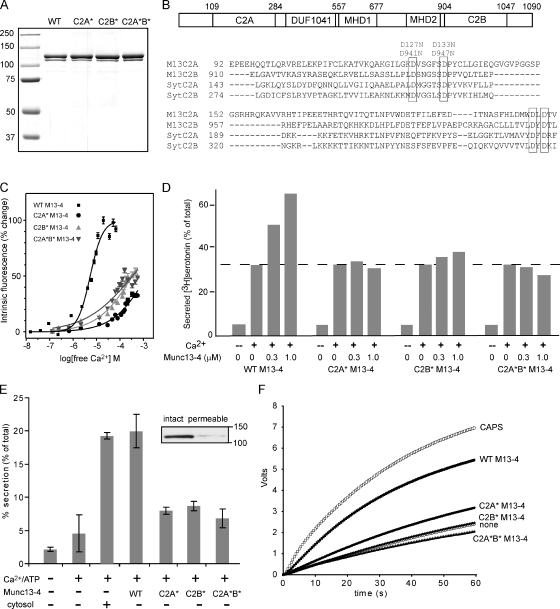
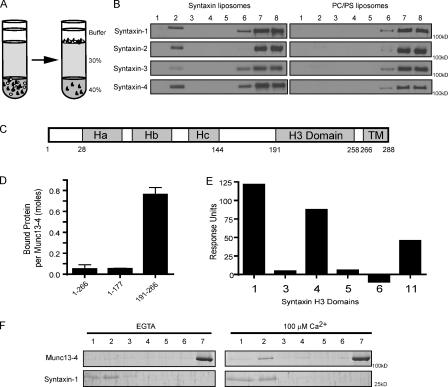
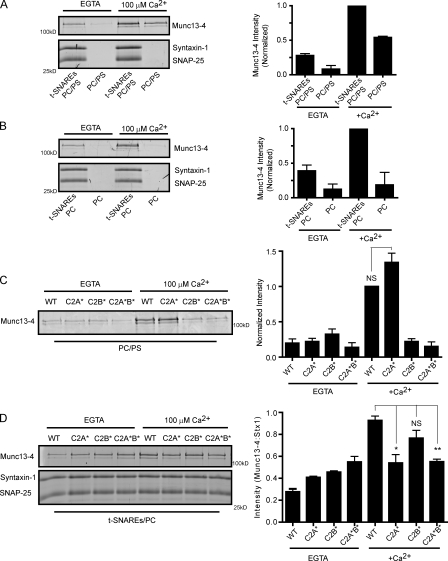
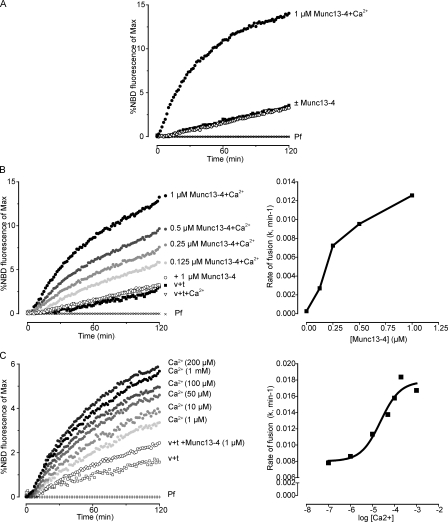
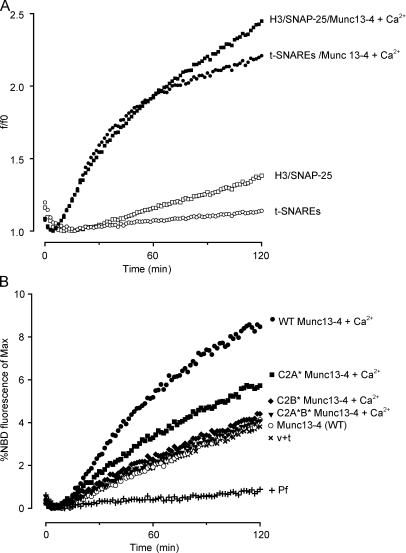
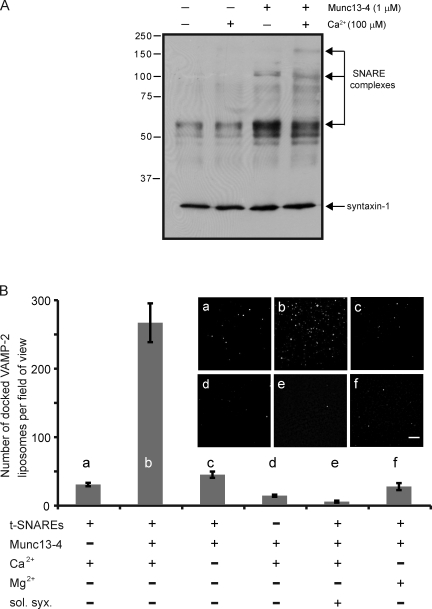
Munc13-4 is a widely expressed member of the CAPS/Munc13 protein family proposed to function in priming secretory granules for exocytosis. Munc13-4 contains N- and C-terminal C2 domains (C2A and C2B) predicted to bind Ca(2+), but Ca(2+)-dependent regulation of Munc13-4 activity has not been described. The C2 domains bracket a predicted SNARE-binding domain, but whether Munc13-4 interacts with SNARE proteins is unknown. We report that Munc13-4 bound Ca(2+) and restored Ca(2+)-dependent granule exocytosis to permeable cells (platelets, mast, and neuroendocrine cells) dependent on putative Ca(2+)-binding residues in C2A and C2B. Munc13-4 exhibited Ca(2+)-stimulated SNARE interactions dependent on C2A and Ca(2+)-dependent membrane binding dependent on C2B. In an apparent coupling of membrane and SNARE binding, Munc13-4 stimulated SNARE-dependent liposome fusion dependent on putative Ca(2+)-binding residues in both C2A and C2B domains. Munc13-4 is the first priming factor shown to promote Ca(2+)-dependent SNARE complex formation and SNARE-mediated liposome fusion. These properties of Munc13-4 suggest its function as a Ca(2+) sensor at rate-limiting priming steps in granule exocytosis.
Figures
References
-
- Arneson L.N., Brickshawana A., Segovis C.M., Schoon R.A., Dick C.J., Leibson P.J. 2007. Cutting edge: Syntaxin 11 regulates lymphocyte-mediated secretion and cytotoxicity. J. Immunol. 179:3397–3401 - PubMed
-
- Bryceson Y.T., Rudd E., Zheng C., Edner J., Ma D., Wood S.M., Bechensteen A.G., Boelens J.J., Celkan T., Farah R.A., et al. 2007. Defective cytotoxic lymphocyte degranulation in syntaxin-11 deficient familial hemophagocytic lymphohistiocytosis 4 (FHL4) patients. Blood. 110:1906–1915 10.1182/blood-2007-02-074468 - DOI - PMC - PubMed
-
- Chen D., Bernstein A.M., Lemons P.P., Whiteheart S.W. 2000. Molecular mechanisms of platelet exocytosis: Role of SNAP-23 and syntaxin 2 in dense core granule release. Blood. 95:921–929 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
