

Membrane binding of Escherichia coli RNase E catalytic domain stabilizes protein structure and increases RNA substrate affinity
- PMID: 22509045
- PMCID: PMC3344982
- DOI: 10.1073/pnas.1120181109
Membrane binding of Escherichia coli RNase E catalytic domain stabilizes protein structure and increases RNA substrate affinity
Abstract
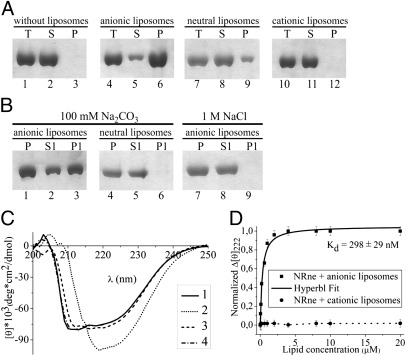
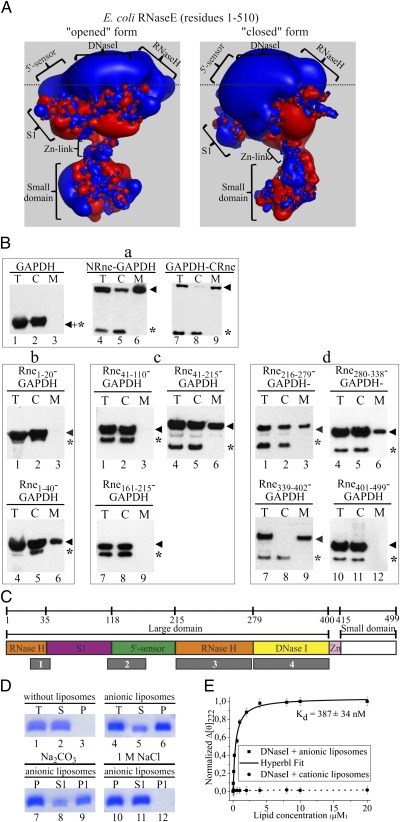
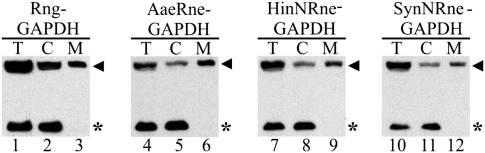
RNase E plays an essential role in RNA processing and decay and tethers to the cytoplasmic membrane in Escherichia coli; however, the function of this membrane-protein interaction has remained unclear. Here, we establish a mechanistic role for the RNase E-membrane interaction. The reconstituted highly conserved N-terminal fragment of RNase E (NRne, residues 1-499) binds specifically to anionic phospholipids through electrostatic interactions. The membrane-binding specificity of NRne was confirmed using circular dichroism difference spectroscopy; the dissociation constant (K(d)) for NRne binding to anionic liposomes was 298 nM. E. coli RNase G and RNase E/G homologs from phylogenetically distant Aquifex aeolicus, Haemophilus influenzae Rd, and Synechocystis sp. were found to be membrane-binding proteins. Electrostatic potentials of NRne and its homologs were found to be conserved, highly positive, and spread over a large surface area encompassing four putative membrane-binding regions identified in the "large" domain (amino acids 1-400, consisting of the RNase H, S1, 5'-sensor, and DNase I subdomains) of E. coli NRne. In vitro cleavage assay using liposome-free and liposome-bound NRne and RNA substrates BR13 and GGG-RNAI showed that NRne membrane binding altered its enzymatic activity. Circular dichroism spectroscopy showed no obvious thermotropic structural changes in membrane-bound NRne between 10 and 60 °C, and membrane-bound NRne retained its normal cleavage activity after cooling. Thus, NRne membrane binding induced changes in secondary protein structure and enzymatic activation by stabilizing the protein-folding state and increasing its binding affinity for its substrate. Our results demonstrate that RNase E-membrane interaction enhances the rate of RNA processing and decay.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
