

A novel intracellular isoform of matrix metalloproteinase-2 induced by oxidative stress activates innate immunity
- PMID: 22509276
- PMCID: PMC3317925
- DOI: 10.1371/journal.pone.0034177
A novel intracellular isoform of matrix metalloproteinase-2 induced by oxidative stress activates innate immunity
Abstract
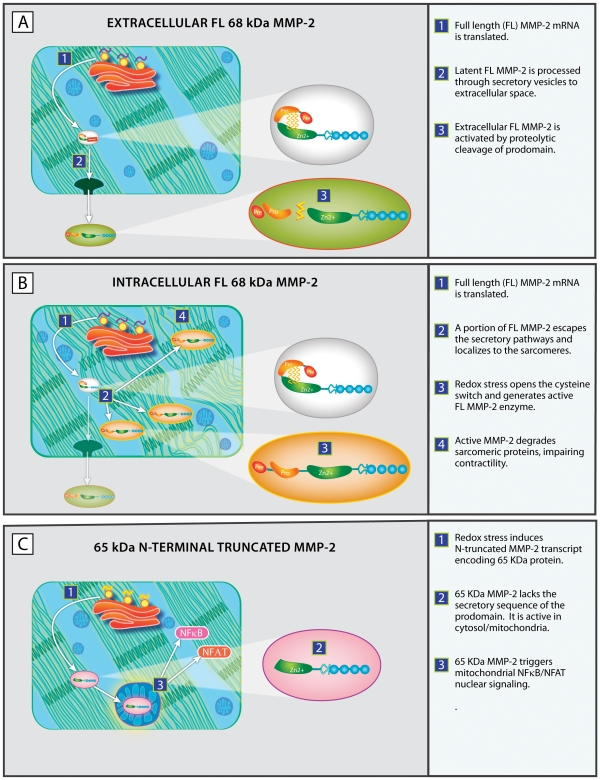
Background: Experimental and clinical evidence has pinpointed a critical role for matrix metalloproteinase-2 (MMP-2) in ischemic ventricular remodeling and systolic heart failure. Prior studies have demonstrated that transgenic expression of the full-length, 68 kDa, secreted form of MMP-2 induces severe systolic failure. These mice also had unexpected and severe mitochondrial structural abnormalities and dysfunction. We hypothesized that an additional intracellular isoform of MMP-2, which affects mitochondrial function is induced under conditions of systolic failure-associated oxidative stress.
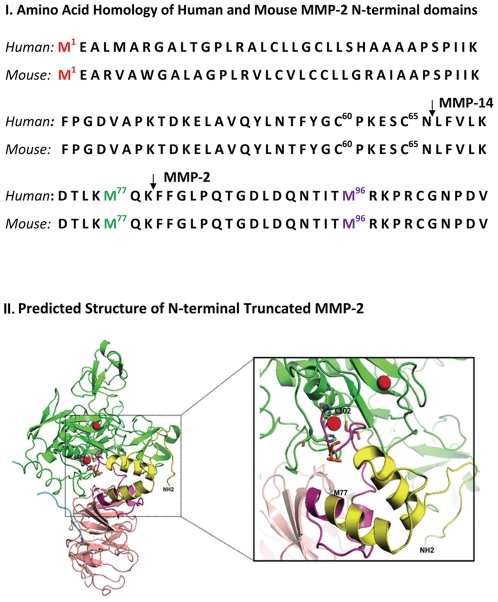
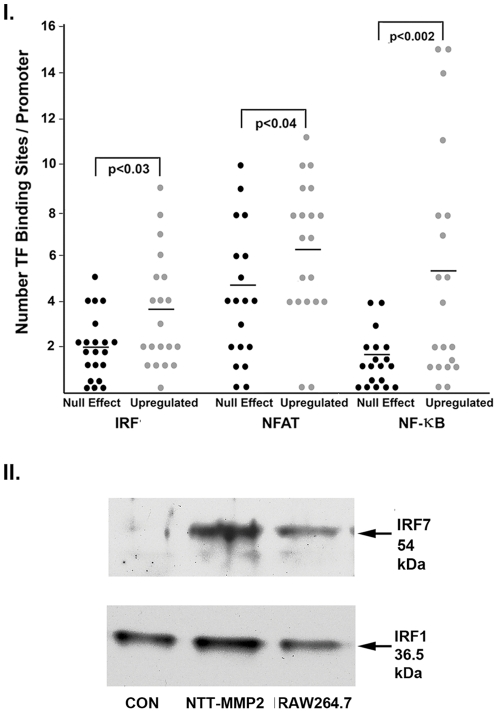
Methodology and principal findings: Western blots of cardiac mitochondria from the full length MMP-2 transgenics, ageing mice and a model of accelerated atherogenesis revealed a smaller 65 kDa MMP-2 isoform. Cultured cardiomyoblasts subjected to transient oxidative stress generated the 65 kDa MMP-2 isoform. The 65 kDa MMP-2 isoform was also induced by hypoxic culture of cardiomyoblasts. Genomic database analysis of the MMP-2 gene mapped transcriptional start sites and RNA transcripts induced by hypoxia or epigenetic modifiers within the first intron of the MMP-2 gene. Translation of these transcripts yields a 65 kDa N-terminal truncated isoform beginning at M(77), thereby deleting the signal sequence and inhibitory prodomain. Cellular trafficking studies demonstrated that the 65 kDa MMP-2 isoform is not secreted and is present in cytosolic and mitochondrial fractions, while the full length 68 kDa isoform was found only in the extracellular space. Expression of the 65 kDa MMP-2 isoform induced mitochondrial-nuclear stress signaling with activation of the pro-inflammatory NF-κB, NFAT and IRF transcriptional pathways. By microarray, the 65 kDa MMP-2 induces an innate immunity transcriptome, including viral stress response genes, innate immunity transcription factor IRF7, chemokines and pro-apoptosis genes.
Conclusion: A novel N-terminal truncated intracellular isoform of MMP-2 is induced by oxidative stress. This isoform initiates a primary innate immune response that may contribute to progressive cardiac dysfunction in the setting of ischemia and systolic failure.
Conflict of interest statement
Figures




Similar articles
-
N-terminal truncated intracellular matrix metalloproteinase-2 induces cardiomyocyte hypertrophy, inflammation and systolic heart failure.PLoS One. 2013 Jul 16;8(7):e68154. doi: 10.1371/journal.pone.0068154. Print 2013. PLoS One. 2013. PMID: 23874529 Free PMC article.
-
An intracellular matrix metalloproteinase-2 isoform induces tubular regulated necrosis: implications for acute kidney injury.Am J Physiol Renal Physiol. 2017 Jun 1;312(6):F1166-F1183. doi: 10.1152/ajprenal.00461.2016. Epub 2017 Mar 22. Am J Physiol Renal Physiol. 2017. PMID: 28331061 Free PMC article.
-
Osteopontin induces nuclear factor kappa B-mediated promatrix metalloproteinase-2 activation through I kappa B alpha /IKK signaling pathways, and curcumin (diferulolylmethane) down-regulates these pathways.J Biol Chem. 2003 Apr 18;278(16):14487-97. doi: 10.1074/jbc.M207309200. Epub 2002 Dec 7. J Biol Chem. 2003. PMID: 12473670
-
Activation of intracellular matrix metalloproteinase-2 by reactive oxygen-nitrogen species: Consequences and therapeutic strategies in the heart.Arch Biochem Biophys. 2013 Dec;540(1-2):82-93. doi: 10.1016/j.abb.2013.09.019. Epub 2013 Oct 5. Arch Biochem Biophys. 2013. PMID: 24103691 Review.
-
Matrix metalloproteinase-2 and myocardial oxidative stress injury: beyond the matrix.Cardiovasc Res. 2010 Feb 1;85(3):413-23. doi: 10.1093/cvr/cvp268. Epub 2009 Aug 4. Cardiovasc Res. 2010. PMID: 19656780 Review.
Cited by
-
Vitamin A, D, E, and K as Matrix Metalloproteinase-2/9 Regulators That Affect Expression and Enzymatic Activity.Int J Mol Sci. 2023 Dec 1;24(23):17038. doi: 10.3390/ijms242317038. Int J Mol Sci. 2023. PMID: 38069361 Free PMC article. Review.
-
Understanding the genetics of intracranial aneurysms: A primer.Clin Neurol Neurosurg. 2022 Jan;212:107060. doi: 10.1016/j.clineuro.2021.107060. Epub 2021 Nov 25. Clin Neurol Neurosurg. 2022. PMID: 34863053 Free PMC article. Review.
-
Matrix Metalloproteinases in Myocardial Infarction and Heart Failure.Prog Mol Biol Transl Sci. 2017;147:75-100. doi: 10.1016/bs.pmbts.2017.02.001. Epub 2017 Mar 18. Prog Mol Biol Transl Sci. 2017. PMID: 28413032 Free PMC article. Review.
-
Time-course RNA-Seq profiling reveals isoform-level gene expression dynamics of the cGAS-STING pathway.Comput Struct Biotechnol J. 2022;20:6490-6500. doi: 10.1016/j.csbj.2022.11.044. Epub 2022 Nov 24. Comput Struct Biotechnol J. 2022. PMID: 36448027 Free PMC article.
-
Matrix Metalloproteinases Repress Hypertrophic Growth in Cardiac Myocytes.Cardiovasc Drugs Ther. 2021 Apr;35(2):353-365. doi: 10.1007/s10557-020-07138-y. Epub 2021 Jan 5. Cardiovasc Drugs Ther. 2021. PMID: 33400052 Free PMC article.
References
-
- Spinale FG. Myocardial matrix remodeling and the matrix metalloproteinases: influence on cardiac form and function. Physiol Rev. 2007;87:1285–1342. - PubMed
-
- Turck J, Pollock AS, Lee LK, Marti H-P, Lovett DH. Matrix metalloproteinase 2 (gelatinase A) regulates glomerular mesangial cell proliferation and differentiation. J Biol Chem. 1996;271:15074–15083. - PubMed
-
- Morgunova E, Tuuttila A, Bergmann U, Isupov M, Lindqvist Y, et al. Structure of human pro-matrix metalloproteinase-2: activation mechanism revealed. Science. 1999;284:1667–70. - PubMed
-
- Coker ML, Doscher MA, Thomas CV, Galis ZS, Spinale FG. Matrix metalloproteinase synthesis and expression in isolated left ventricular myocyte preparation. Amer J Physiol Heart Circ Physiol. 1999;277:H777–787. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
