

Phosphorylation of Ubc9 by Cdk1 enhances SUMOylation activity
- PMID: 22509284
- PMCID: PMC3317942
- DOI: 10.1371/journal.pone.0034250
Phosphorylation of Ubc9 by Cdk1 enhances SUMOylation activity
Abstract
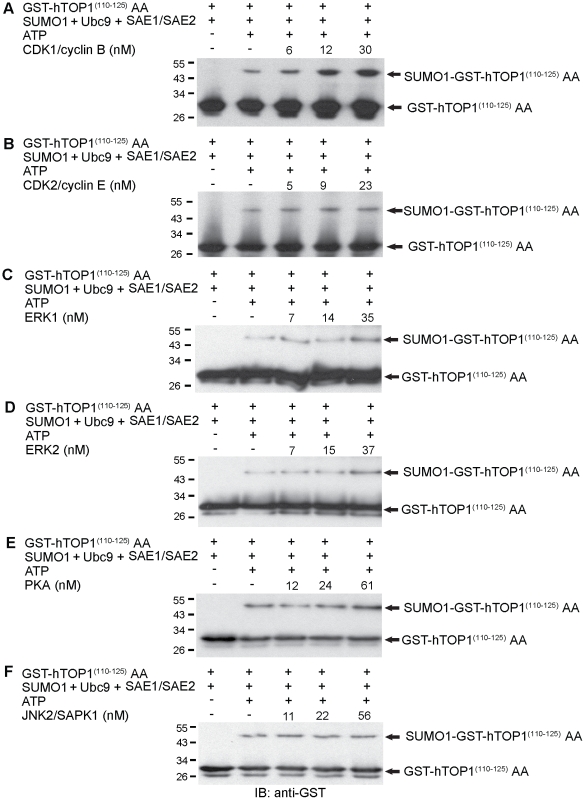
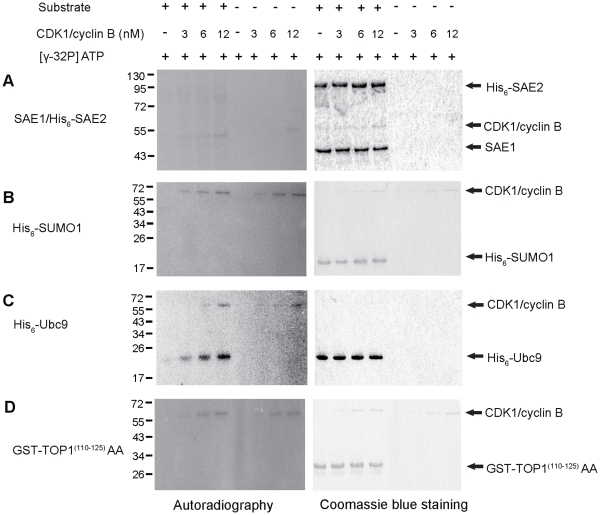
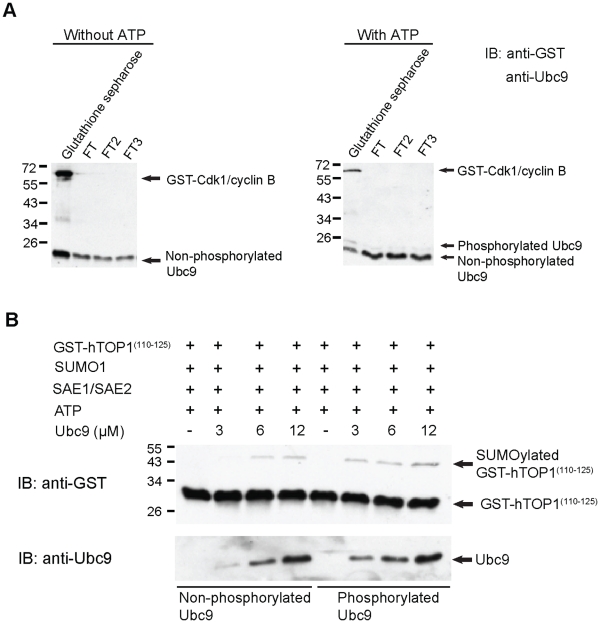
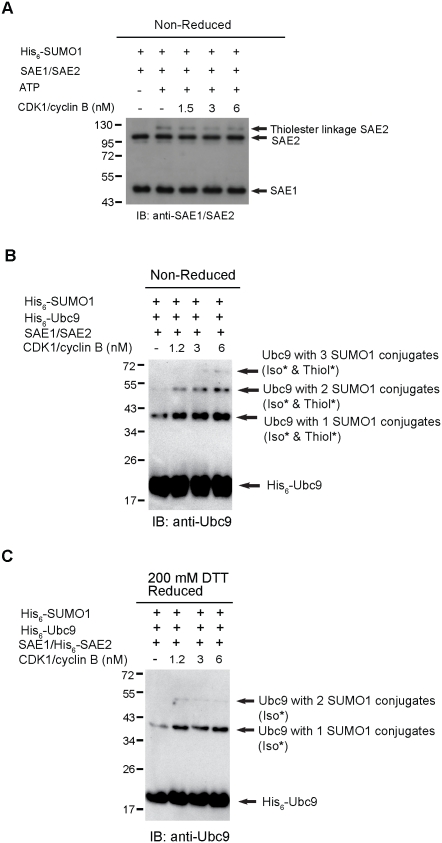
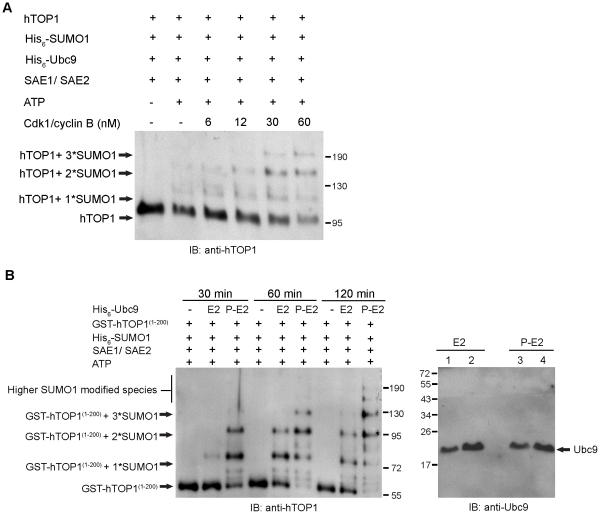
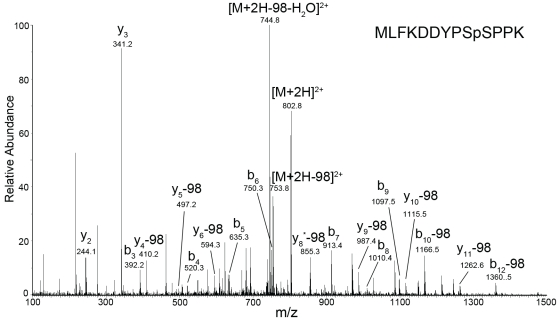
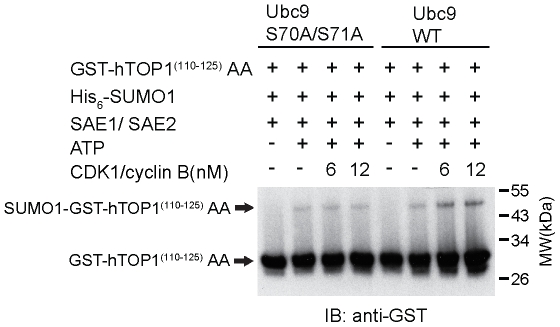
Increasing evidence has pointed to an important role of SUMOylation in cell cycle regulation, especially for M phase. In the current studies, we have obtained evidence through in vitro studies that the master M phase regulator CDK1/cyclin B kinase phosphorylates the SUMOylation machinery component Ubc9, leading to its enhanced SUMOylation activity. First, we show that CDK1/cyclin B, but not many other cell cycle kinases such as CDK2/cyclin E, ERK1, ERK2, PKA and JNK2/SAPK1, specifically enhances SUMOylation activity. Second, CDK1/cyclin B phosphorylates the SUMOylation machinery component Ubc9, but not SAE1/SAE2 or SUMO1. Third, CDK1/cyclin B-phosphorylated Ubc9 exhibits increased SUMOylation activity and elevated accumulation of the Ubc9-SUMO1 thioester conjugate. Fourth, CDK1/cyclin B enhances SUMOylation activity through phosphorylation of Ubc9 at serine 71. These studies demonstrate for the first time that the cell cycle-specific kinase CDK1/cyclin B phosphorylates a SUMOylation machinery component to increase its overall SUMOylation activity, suggesting that SUMOylation is part of the cell cycle program orchestrated by CDK1 through Ubc9.
Conflict of interest statement
Figures
References
-
- Hay RT. SUMO: a history of modification. Mol Cell. 2005;18:1–12. - PubMed
-
- Seufert W, Futcher B, Jentsch S. Role of a ubiquitin-conjugating enzyme in degradation of S- and M-phase cyclins. Nature. 1995;373:78–81. - PubMed
-
- al-Khodairy F, Enoch T, Hagan IM, Carr AM. The Schizosaccharomyces pombe hus5 gene encodes a ubiquitin conjugating enzyme required for normal mitosis. J Cell Sci 108 ( Pt. 1995;2):475–486. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
