

Corticotropin-releasing factor acting at the locus coeruleus disrupts thalamic and cortical sensory-evoked responses
- PMID: 22510725
- PMCID: PMC3398725
- DOI: 10.1038/npp.2012.50
Corticotropin-releasing factor acting at the locus coeruleus disrupts thalamic and cortical sensory-evoked responses
Abstract
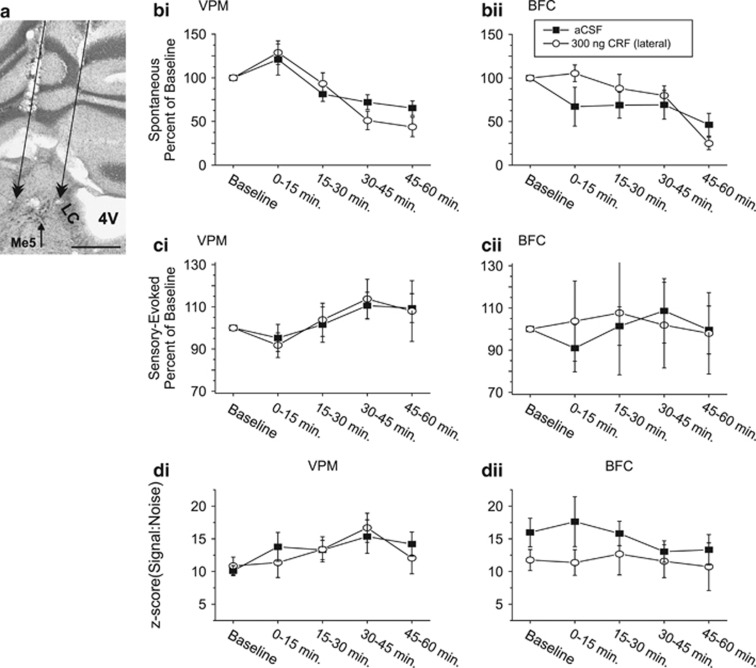
Stress and stress-related psychiatric disorders, including post-traumatic stress disorder, are associated with disruptions in sensory information processing. The neuropeptide, corticotropin-releasing factor (CRF), coordinates the physiological and behavioral responses to stress, in part, by activating the locus coeruleus-norepinephrine (LC-NE) projection system. Although the LC-NE system is an important modulator of sensory information processing, to date, the consequences of CRF activation of this system on sensory signal processing are poorly understood. The current study examined the dose-dependent actions of CRF at the LC on spontaneous and sensory-evoked discharge of neurons within the thalamus and cortex of the vibrissa somatosensory system in the awake, freely moving rat. Peri-LC infusions of CRF resulted in a dose-dependent suppression of sensory-evoked discharge in ventral posterior medial thalamic and barrel field cortical neurons. A concurrent increase in spontaneous activity was observed. This latter action is generally not found with iontophoretic application of NE to target neurons or stimulation of the LC-NE pathway. Net decreases in signal-to-noise of sensory-evoked responses within both regions suggest that under conditions associated with CRF release at the LC, including stress, the transfer of afferent information within sensory systems is impaired. Acutely, a suppression of certain types of sensory information may represent an adaptive response to an immediate unexpected stressor. Persistence of such effects could contribute to abnormalities of information processing seen in sensorimotor gating associated with stress and stress-related psychopathology.
Figures




Similar articles
-
The effects of tonic locus ceruleus output on sensory-evoked responses of ventral posterior medial thalamic and barrel field cortical neurons in the awake rat.J Neurosci. 2004 Dec 1;24(48):10773-85. doi: 10.1523/JNEUROSCI.1573-04.2004. J Neurosci. 2004. PMID: 15574728 Free PMC article.
-
Effects of intracerebroventricular corticotropin releasing factor on sensory-evoked responses in the rat visual thalamus.Brain Res. 2014 May 2;1561:35-47. doi: 10.1016/j.brainres.2014.02.048. Epub 2014 Mar 22. Brain Res. 2014. PMID: 24661913 Free PMC article.
-
Discrete local application of corticotropin-releasing factor increases locus coeruleus discharge and extracellular norepinephrine in rat hippocampus.Synapse. 1999 Sep 15;33(4):304-13. doi: 10.1002/(SICI)1098-2396(19990915)33:4<304::AID-SYN7>3.0.CO;2-Z. Synapse. 1999. PMID: 10421711
-
Pharmacology of locus coeruleus spontaneous and sensory-evoked activity.Prog Brain Res. 1991;88:249-56. doi: 10.1016/s0079-6123(08)63814-5. Prog Brain Res. 1991. PMID: 1813924 Review.
-
Opposing regulation of the locus coeruleus by corticotropin-releasing factor and opioids. Potential for reciprocal interactions between stress and opioid sensitivity.Psychopharmacology (Berl). 2001 Dec;158(4):331-42. doi: 10.1007/s002130000673. Epub 2001 Mar 14. Psychopharmacology (Berl). 2001. PMID: 11797054 Review.
Cited by
-
Xiao Yao San Improves Depressive-Like Behaviors in Rats with Chronic Immobilization Stress through Modulation of Locus Coeruleus-Norepinephrine System.Evid Based Complement Alternat Med. 2014;2014:605914. doi: 10.1155/2014/605914. Epub 2014 Dec 25. Evid Based Complement Alternat Med. 2014. PMID: 25610478 Free PMC article.
-
Locus coeruleus kappa-opioid receptors modulate reinstatement of cocaine place preference through a noradrenergic mechanism.Neuropsychopharmacology. 2013 Nov;38(12):2484-97. doi: 10.1038/npp.2013.151. Epub 2013 Jun 21. Neuropsychopharmacology. 2013. PMID: 23787819 Free PMC article.
-
Region-specific roles of the corticotropin-releasing factor-urocortin system in stress.Nat Rev Neurosci. 2016 Oct;17(10):636-51. doi: 10.1038/nrn.2016.94. Epub 2016 Sep 2. Nat Rev Neurosci. 2016. PMID: 27586075 Review.
-
The role of the dynorphin/κ opioid receptor system in anxiety.Acta Pharmacol Sin. 2015 Jul;36(7):783-90. doi: 10.1038/aps.2015.32. Epub 2015 May 18. Acta Pharmacol Sin. 2015. PMID: 25982631 Free PMC article. Review.
-
Ketamine influences the locus coeruleus norepinephrine network, with a dependency on norepinephrine transporter genotype - a placebo controlled fMRI study.Neuroimage Clin. 2018;20:715-723. doi: 10.1016/j.nicl.2018.09.001. Epub 2018 Sep 4. Neuroimage Clin. 2018. PMID: 30238915 Free PMC article. Clinical Trial.
References
-
- Bale TL, Vale WW. CRF and CRF receptors: role in stress responsivity and other behaviors. Annu Rev Pharmacol Toxicol. 2004;44:525–557. - PubMed
-
- Barrios-Choplin B, McCraty R, Cryer B. An inner quality approach to reducing stress and improving physical and emotional wellbeing at work. Stress Medicine. 1998;13:193–201.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
