

Mechanisms of tissue fusion during development
- PMID: 22510983
- PMCID: PMC3328173
- DOI: 10.1242/dev.068338
Mechanisms of tissue fusion during development
Abstract
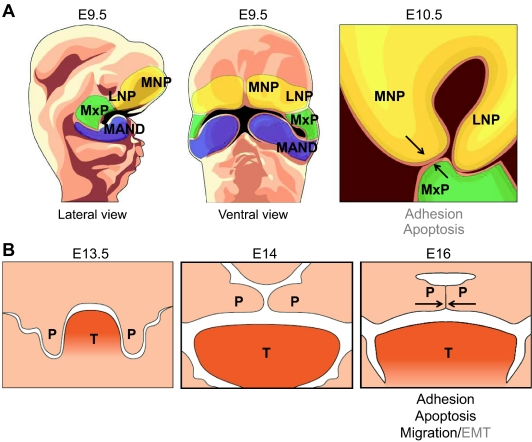
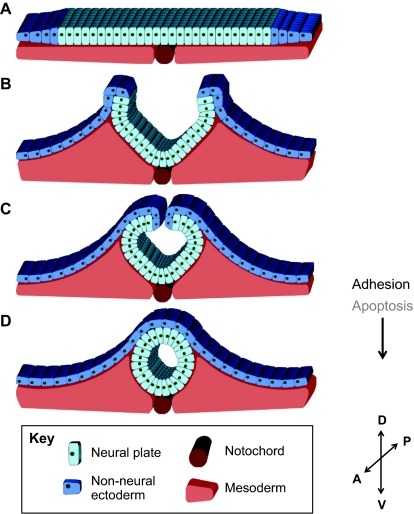
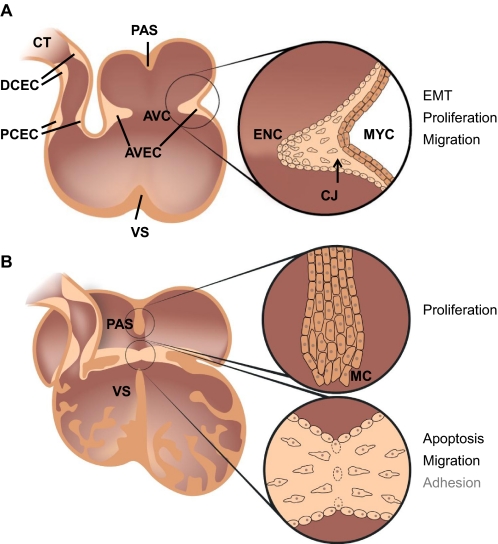
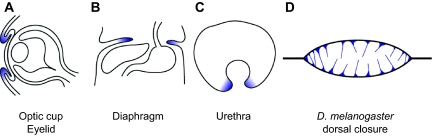
Tissue fusion events during embryonic development are crucial for the correct formation and function of many organs and tissues, including the heart, neural tube, eyes, face and body wall. During tissue fusion, two opposing tissue components approach one another and integrate to form a continuous tissue; disruption of this process leads to a variety of human birth defects. Genetic studies, together with recent advances in the ability to culture developing tissues, have greatly enriched our knowledge of the mechanisms involved in tissue fusion. This review aims to bring together what is currently known about tissue fusion in several developing mammalian organs and highlights some of the questions that remain to be addressed.
Figures
References
-
- Abbott B. D. (2010). The etiology of cleft palate: a 50-year search for mechanistic and molecular understanding. Birth Defects Res. B Dev. Reprod. Toxicol. 89, 266–274 - PubMed
-
- Abdul-Aziz N. M., Turmaine M., Greene N. D., Copp A. J. (2009). EphrinA-EphA receptor interactions in mouse spinal neurulation: implications for neural fold fusion. Int. J. Dev. Biol. 53, 559–568 - PubMed
-
- Beverdam A., Brouwer A., Reijnen M., Korving J., Meijlink F. (2001). Severe nasal clefting and abnormal embryonic apoptosis in Alx3/Alx4 double mutant mice. Development 128, 3975–3986 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
