

Functional interactions between Fat family cadherins in tissue morphogenesis and planar polarity
- PMID: 22510986
- PMCID: PMC3328180
- DOI: 10.1242/dev.077461
Functional interactions between Fat family cadherins in tissue morphogenesis and planar polarity
Abstract
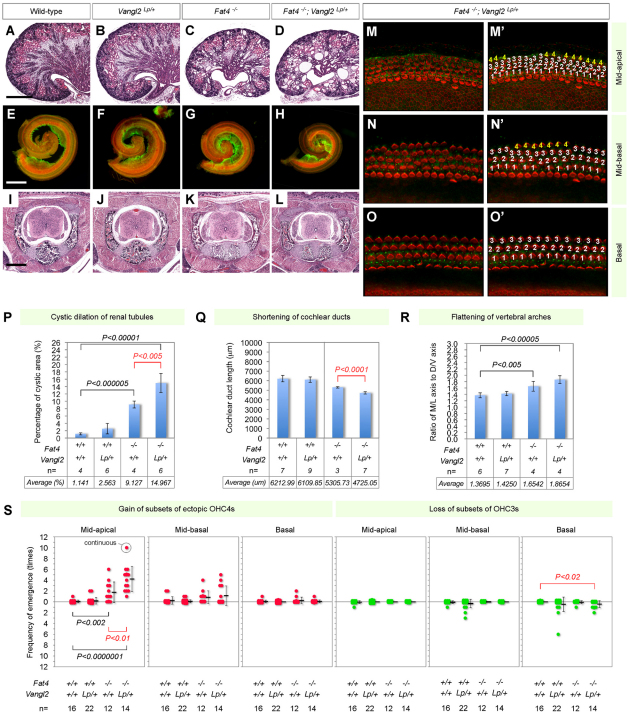
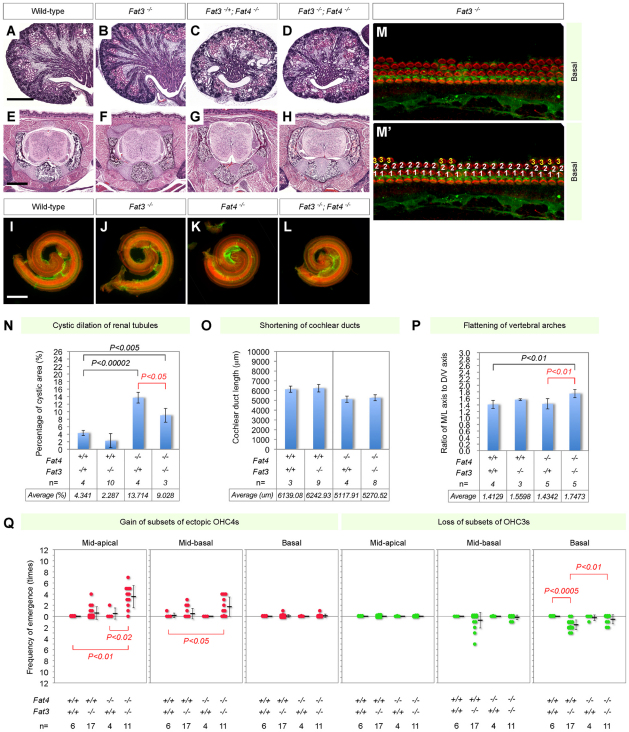
The atypical cadherin fat (ft) was originally discovered as a tumor suppressor in Drosophila and later shown to regulate a form of tissue patterning known as planar polarity. In mammals, four ft homologs have been identified (Fat1-4). Recently, we demonstrated that Fat4 plays a role in vertebrate planar polarity. Fat4 has the highest homology to ft, whereas other Fat family members are homologous to the second ft-like gene, ft2. Genetic studies in flies and mice imply significant functional differences between the two groups of Fat cadherins. Here, we demonstrate that Fat family proteins act both synergistically and antagonistically to influence multiple aspects of tissue morphogenesis. We find that Fat1 and Fat4 cooperate during mouse development to control renal tubular elongation, cochlear extension, cranial neural tube formation and patterning of outer hair cells in the cochlea. Similarly, Fat3 and Fat4 synergize to drive vertebral arch fusion at the dorsal midline during caudal vertebra morphogenesis. We provide evidence that these effects depend on conserved interactions with planar polarity signaling components. In flies, the transcriptional co-repressor Atrophin (Atro) physically interacts with Ft and acts as a component of Fat signaling for planar polarity. We find that the mammalian orthologs of atro, Atn1 and Atn2l, modulate Fat4 activity during vertebral arch fusion and renal tubular elongation, respectively. Moreover, Fat4 morphogenetic defects are enhanced by mutations in Vangl2, a 'core' planar cell polarity gene. These studies highlight the wide range and complexity of Fat activities and suggest that a Fat-Atrophin interaction is a conserved element of planar polarity signaling.
Figures






References
-
- Axelrod J. D. (2009). Progress and challenges in understanding planar cell polarity signaling. Semin. Cell Dev. Biol. 20, 964–971 - PubMed
-
- Banting G. S., Barak O., Ames T. M., Burnham A. C., Kardel M. D., Cooch N. S., Davidson C. E., Godbout R., McDermid H. E., Shiekhattar R. (2005). CECR2, a protein involved in neurulation, forms a novel chromatin remodeling complex with SNF2L. Hum. Mol. Genet. 14, 513–524 - PubMed
-
- Brooker R., Hozumi K., Lewis J. (2006). Notch ligands with contrasting functions: Jagged1 and Delta1 in the mouse inner ear. Development 133, 1277–1286 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
