

BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles
- PMID: 22511774
- PMCID: PMC3365991
- DOI: 10.1074/jbc.M112.351908
BLOC-2, AP-3, and AP-1 proteins function in concert with Rab38 and Rab32 proteins to mediate protein trafficking to lysosome-related organelles
Abstract
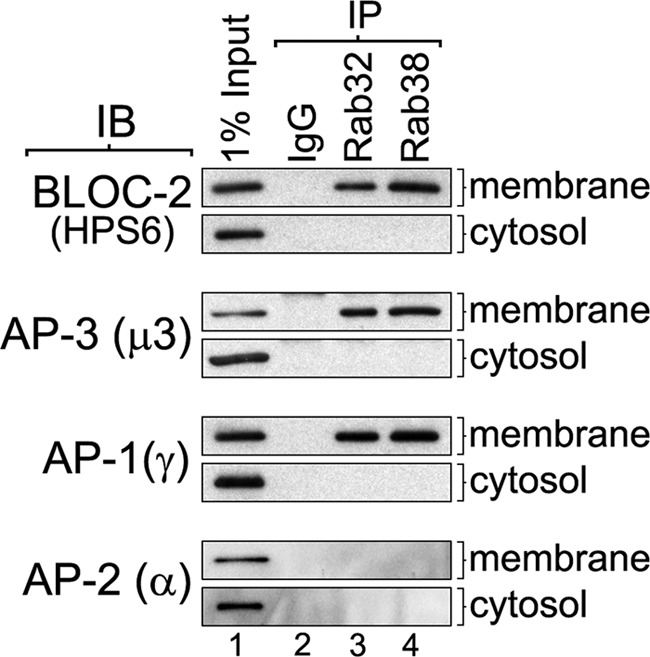
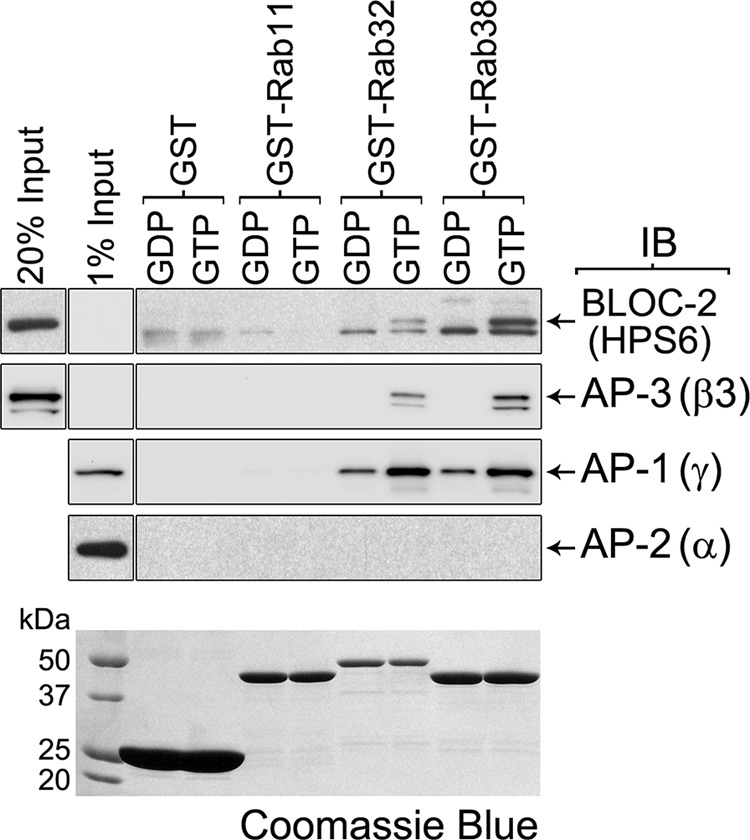
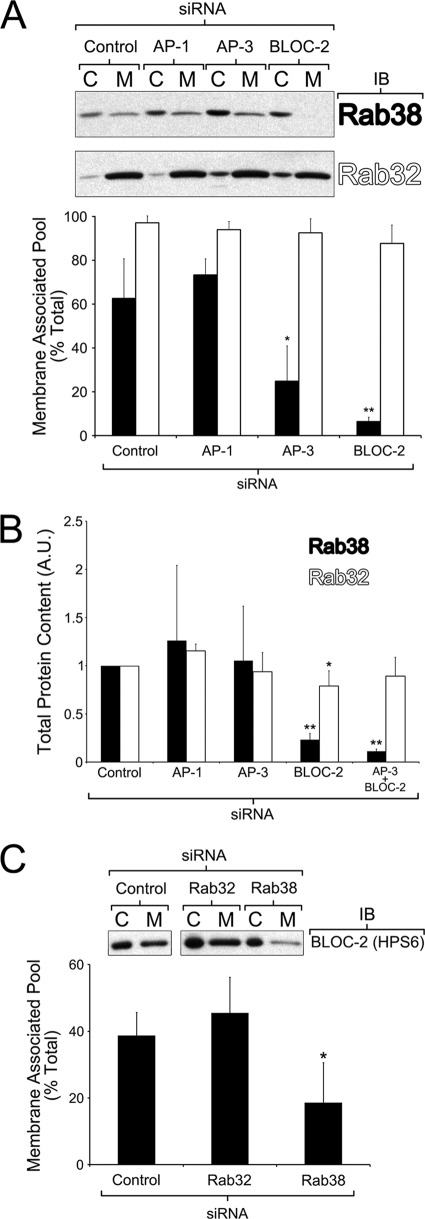
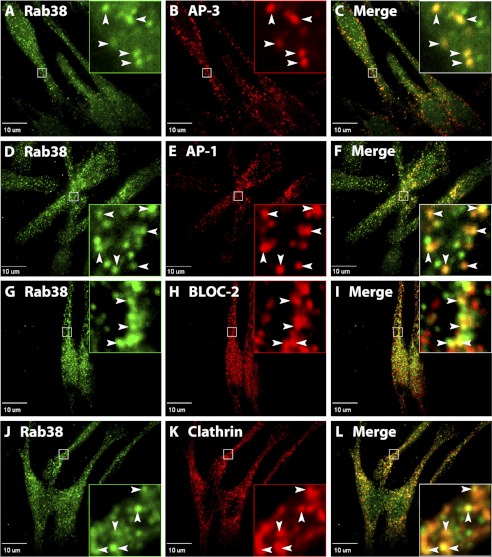
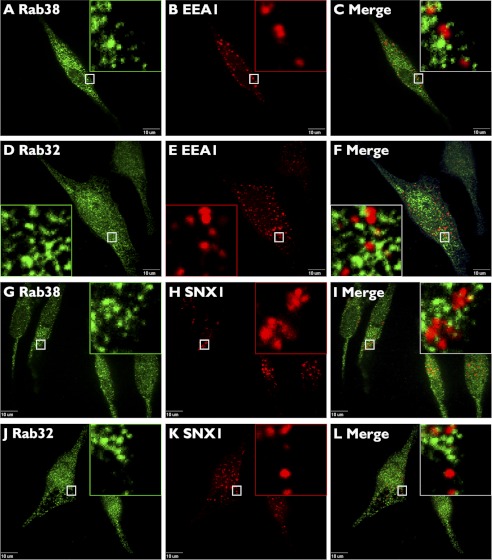
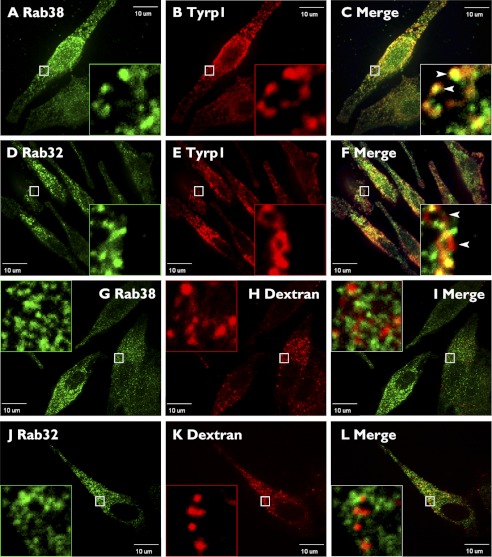
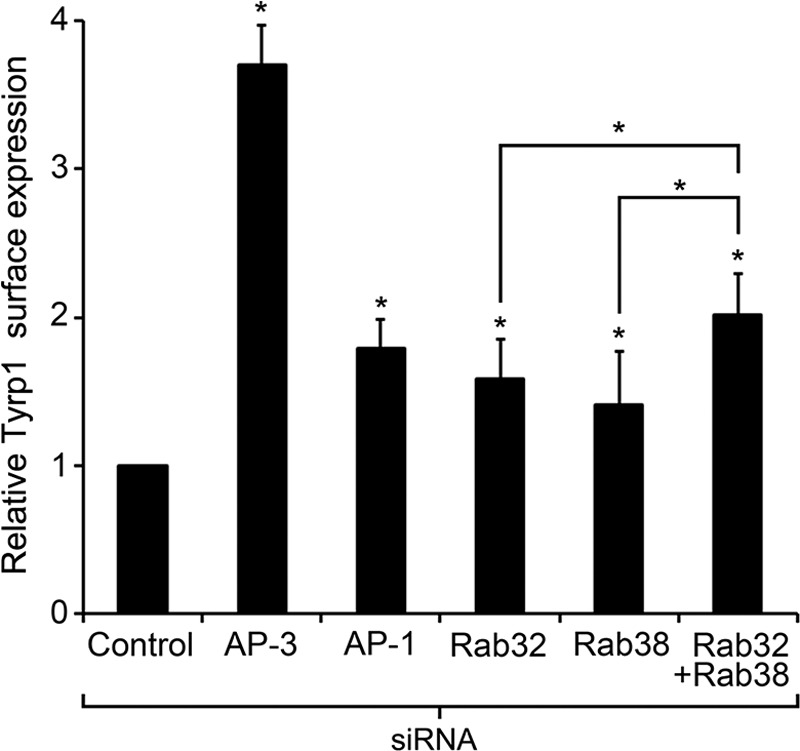
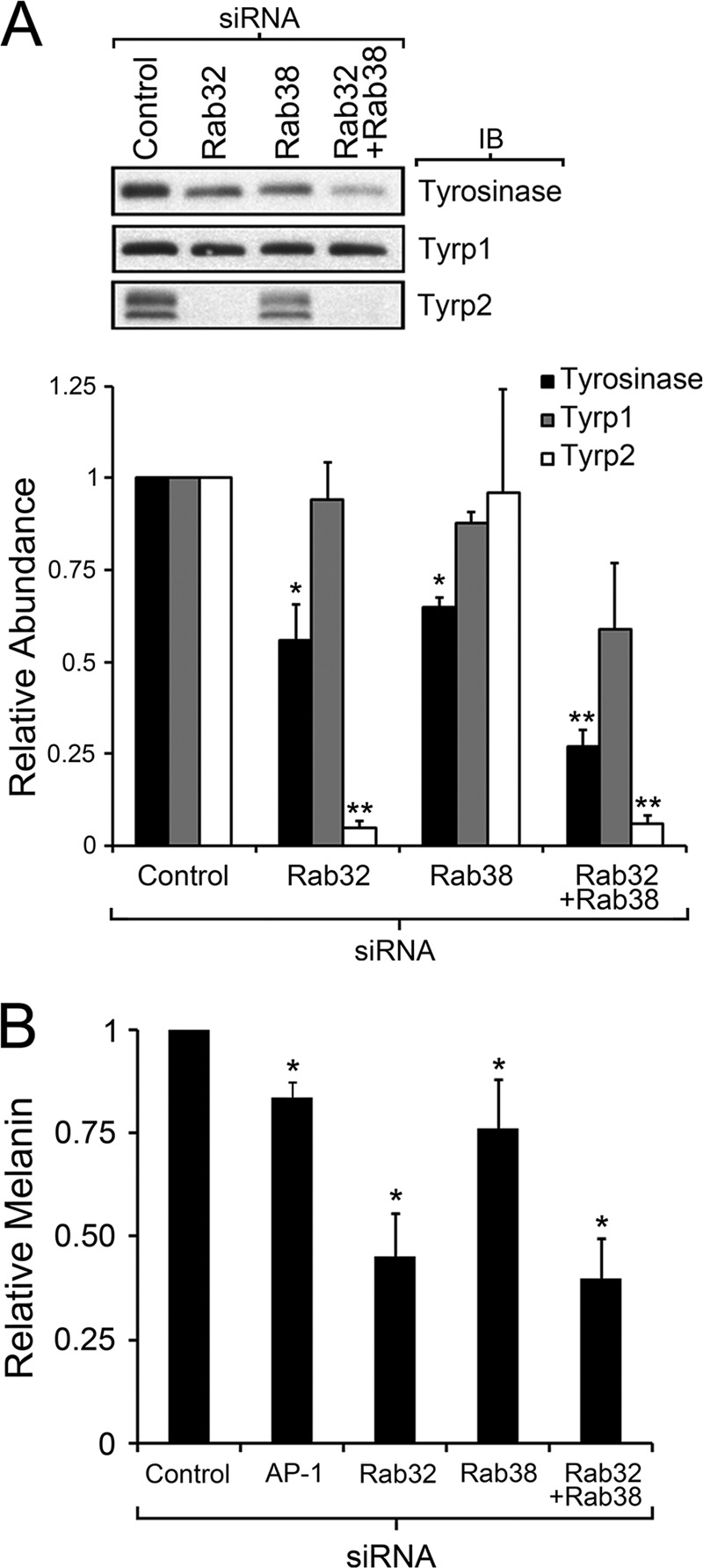
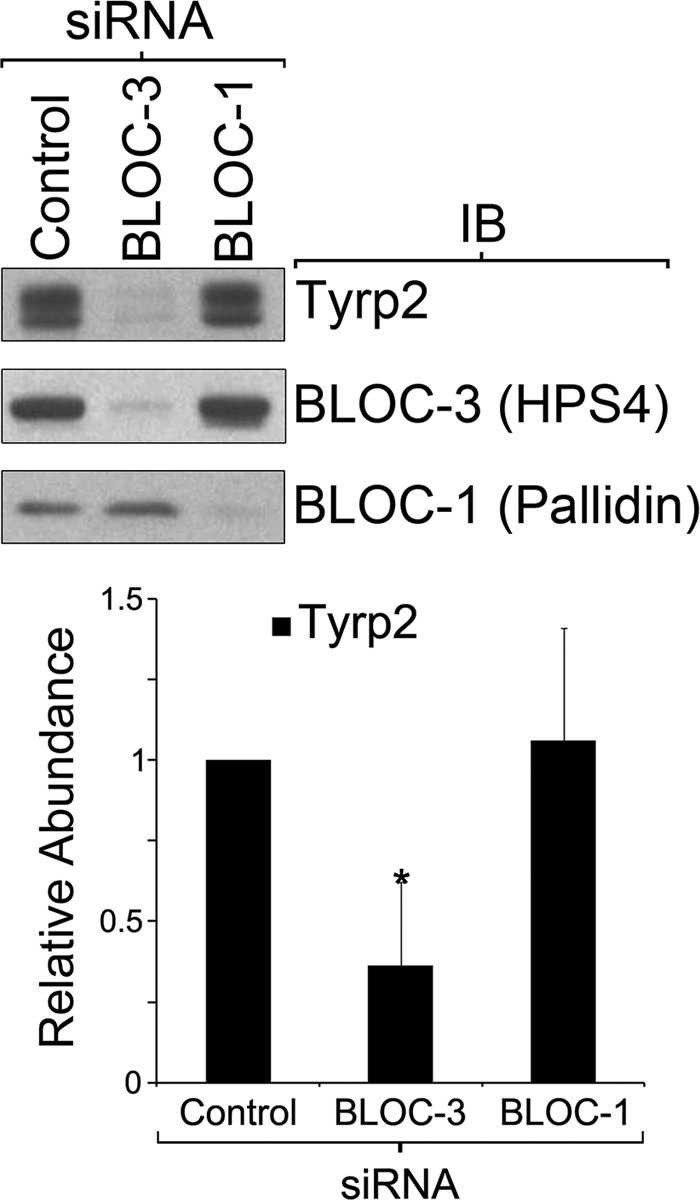
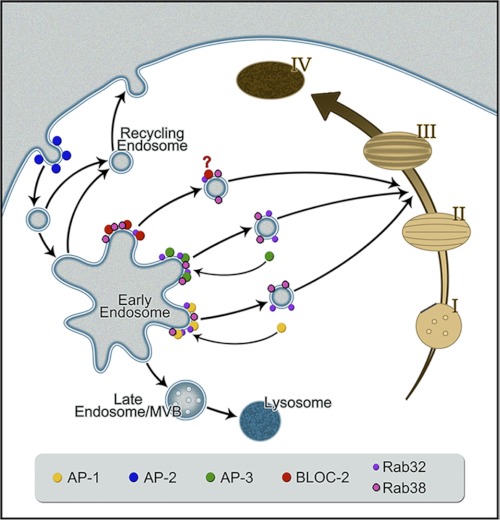
Lysosome-related organelles (LROs) are synthesized in specialized cell types where they largely coexist with conventional lysosomes. Most of the known cellular transport machinery involved in biogenesis are ubiquitously expressed and shared between lysosomes and LROs. Examples of common components are the adaptor protein complex-3 (AP-3) and biogenesis of lysosome-related organelle complex (BLOC)-2. These protein complexes control sorting and transport of newly synthesized integral membrane proteins from early endosomes to both lysosomes and LROs such as the melanosome. However, it is unknown what factors cooperate with the ubiquitous transport machinery to mediate transport to LROs in specialized cells. Focusing on the melanosome, we show that the ubiquitous machinery interacts with cell type-specific Rab proteins, Rab38 and Rab32, to facilitate transport to the maturing organelle. BLOC-2, AP-3, and AP-1 coimmunoprecipitated with Rab38 and Rab32 from MNT-1 melanocytic cell extracts. BLOC-2, AP-3, AP-1, and clathrin partially colocalized with Rab38 and Rab32 by confocal immunofluorescence microscopy in MNT-1 cells. Rab38- and Rab32-deficient MNT-1 cells displayed abnormal trafficking and steady state levels of known cargoes of the BLOC-2, AP-3, and AP-1 pathways, the melanin-synthesizing enzymes tyrosinase and tyrosinase-related protein-1. These observations support the idea that Rab38 and Rab32 are the specific factors that direct the ubiquitous machinery to mediate transport from early endosomes to maturing LROs. Additionally, analysis of tyrosinase-related protein-2 and total melanin production indicates that Rab32 has unique functions that cannot be carried out by Rab38 in melanosome biogenesis.
Figures
References
-
- Dell'Angelica E. C., Mullins C., Caplan S., Bonifacino J. S. (2000) Lysosome-related organelles. FASEB J. 14, 1265–1278 - PubMed
-
- Spritz R. A., Chiang P. W., Oiso N., Alkhateeb A. (2003) Human and mouse disorders of pigmentation. Curr. Opin. Genet. Dev. 13, 284–289 - PubMed
-
- Di Pietro S. M., Dell'Angelica E. C. (2005) The cell biology of Hermansky-Pudlak syndrome. Recent advances. Traffic 6, 525–533 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
