

Understanding tinnitus: the dorsal cochlear nucleus, organization and plasticity
- PMID: 22513100
- PMCID: PMC3402636
- DOI: 10.1016/j.brainres.2012.03.044
Understanding tinnitus: the dorsal cochlear nucleus, organization and plasticity
Abstract
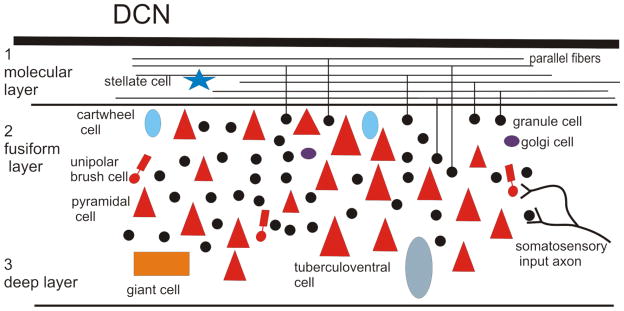
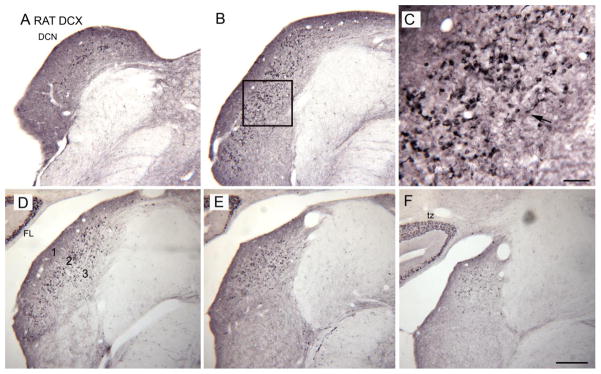
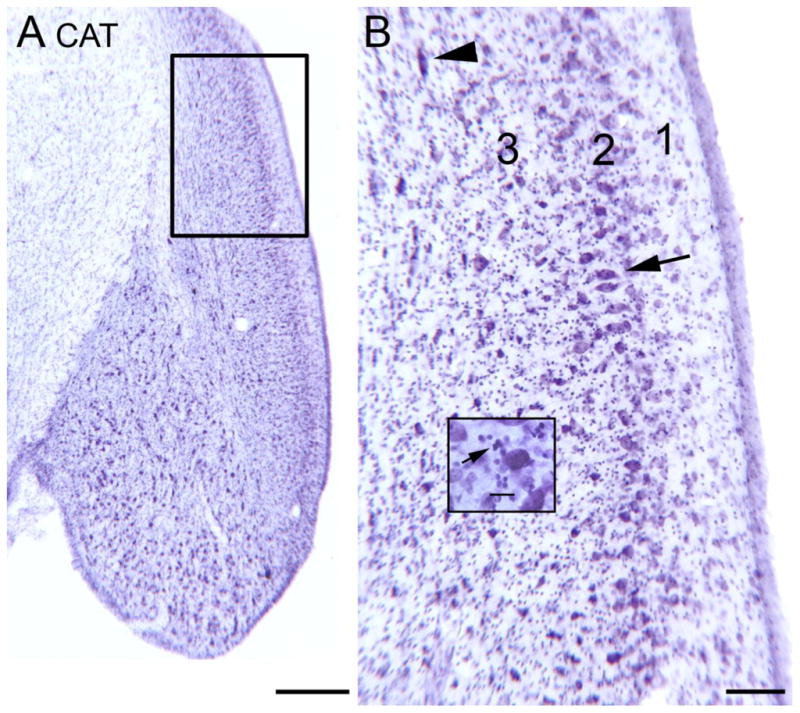
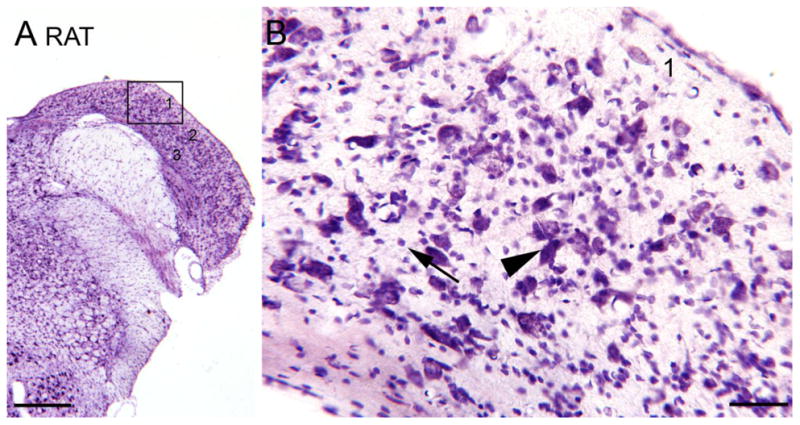
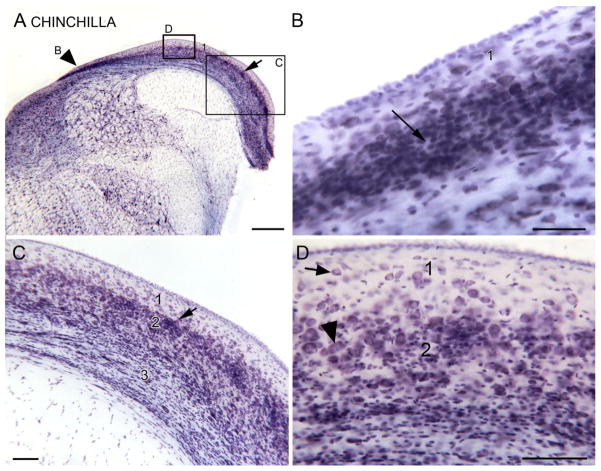
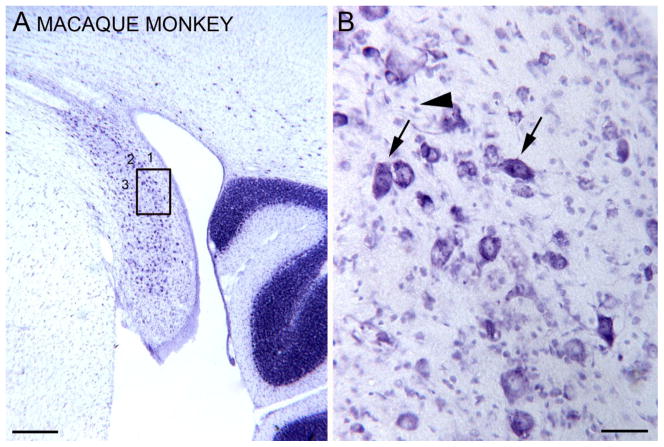
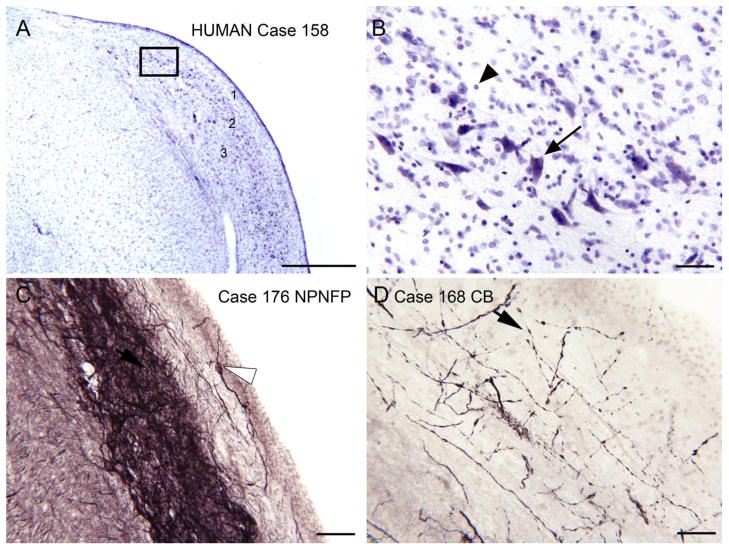
Tinnitus, the perception of a phantom sound, is a common consequence of damage to the auditory periphery. A major goal of tinnitus research is to find the loci of the neural changes that underlie the disorder. Crucial to this endeavor has been the development of an animal behavioral model of tinnitus, so that neural changes can be correlated with behavioral evidence of tinnitus. Three major lines of evidence implicate the dorsal cochlear nucleus (DCN) in tinnitus. First, elevated spontaneous activity in the DCN is correlated with peripheral damage and tinnitus. Second, there are somatosensory inputs to the DCN that can modulate spontaneous activity and might mediate the somatic-auditory interactions seen in tinnitus patients. Third, we have found a subpopulation of DCN neurons in the adult rat that express doublecortin, a plasticity-related protein. The expression of this protein may reflect a role of these neurons in the neural reorganization causing tinnitus. However, there is a problem in extending the findings in the rodent DCN to humans. Classic studies state that the structure of the primate DCN is quite different from that of rodents, with primates lacking granule cells, the recipients of somatosensory input. To address the possibility of major species differences in DCN organization, we compared Nissl-stained sections of the DCN in five different species. In contrast to earlier reports, our data suggest that the organization of the primate DCN is not dramatically different from that of the rodents, and validate the use of animal data in the study of tinnitus. This article is part of a Special Issue entitled: Tinnitus Neuroscience.
Copyright © 2012 Elsevier B.V. All rights reserved.
Figures
References
-
- Abbott SD, Hughes LF, Bauer CA, Salvi R, Caspary DM. Detection of glutamate decarboxylase isoforms in rat inferior colliculus following acoustic exposure. Neuroscience. 1999;93:1375–81. - PubMed
-
- Adams JC. Neuronal morphology in the human cochlear nucleus. Arch Otolaryngol Head Neck Surg. 1986;112:1253–61. - PubMed
-
- Adjamian P, Sereda M, Hall DA. The mechanisms of tinnitus: perspectives from human functional neuroimaging. Hear Res. 2009;253:15–31. - PubMed
-
- Aitkin LM, Phillips SC. Is the inferior colliculus an obligatory relay in the cat auditory system? Neurosci Lett. 1984;44:259–64. - PubMed
-
- Aitkin LM, Pettigrew JD, Calford MB, Phillips SC, Wise LZ. Representation of stimulus azimuth by low-frequency neurons in inferior colliculus of the cat. J Neurophysiol. 1985;53:43–59. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
