KCNE2 and the K (+) channel: the tail wagging the dog
- PMID: 22513486
- PMCID: PMC3367673
- DOI: 10.4161/chan.19126
KCNE2 and the K (+) channel: the tail wagging the dog
Abstract
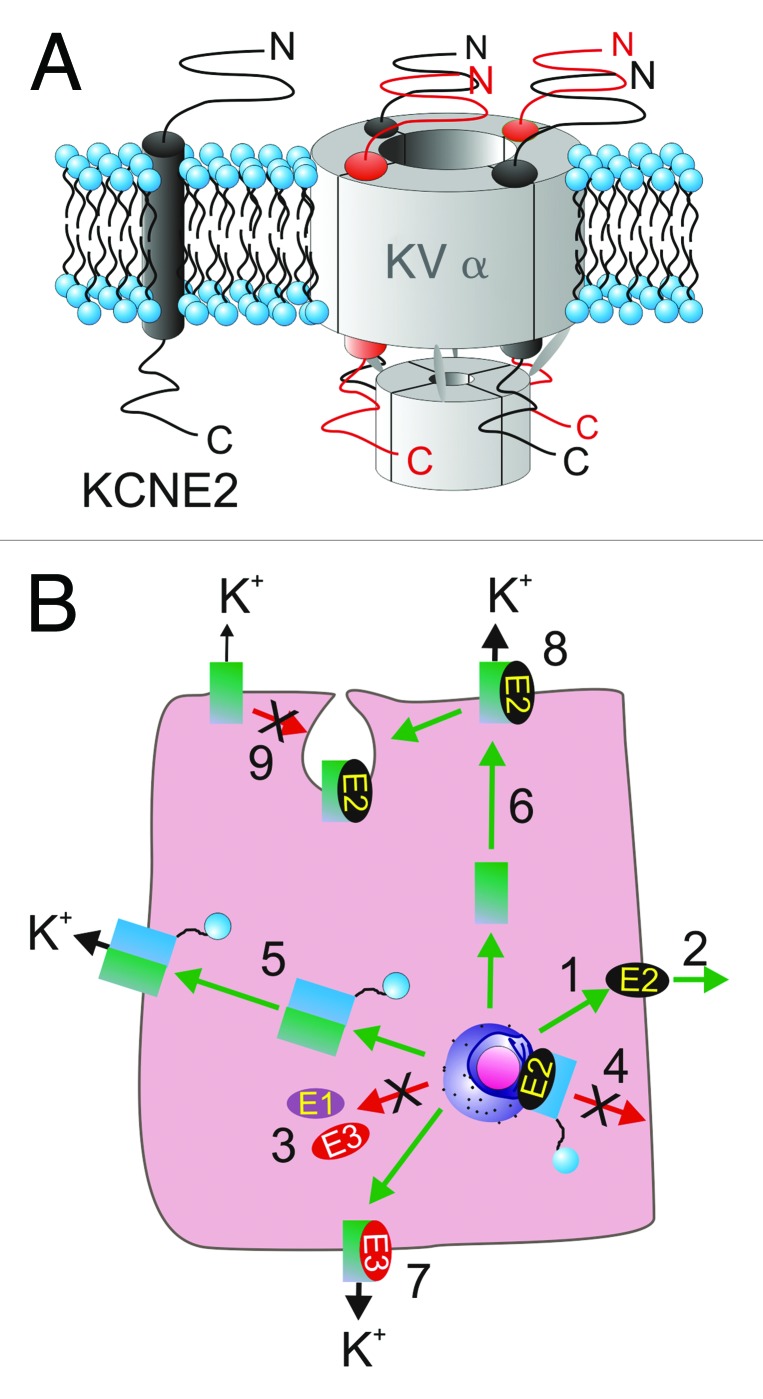
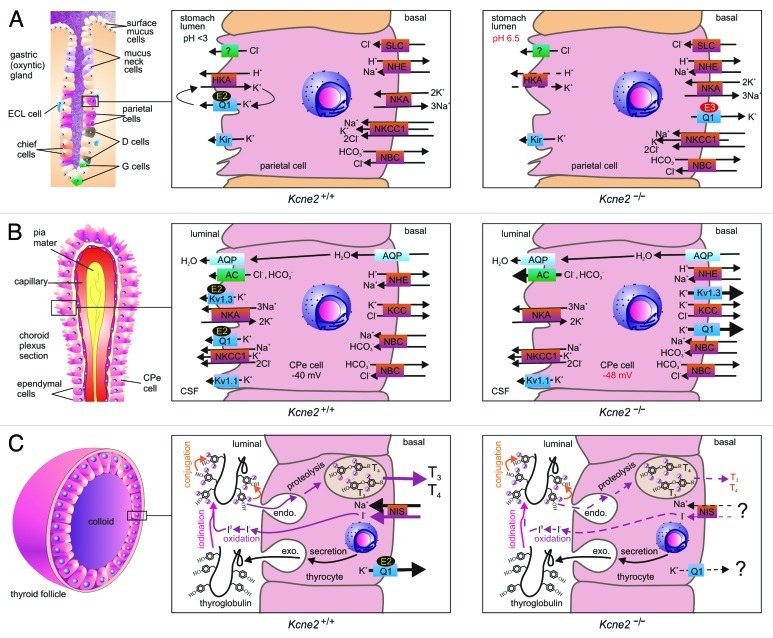
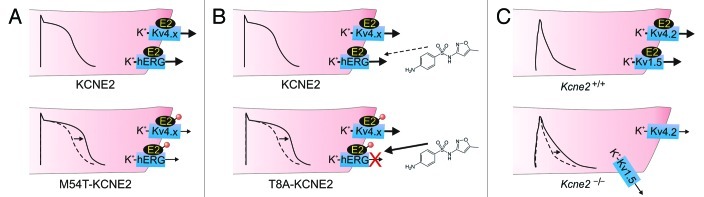
KCNE2, originally designated MinK-related peptide 1 (MiRP1), belongs to a five-strong family of potassium channel ancillary (β) subunits that, despite the diminutive size of the family and its members, has loomed large in the field of ion channel physiology. KCNE2 dictates K (+) channel gating, conductance, α subunit composition, trafficking and pharmacology, and also modifies functional properties of monovalent cation-nonselective HCN channels. The Kcne2 (-/-) mouse exhibits cardiac arrhythmia and hypertrophy, achlorhydria, gastric neoplasia, hypothyroidism, alopecia, stunted growth and choroid plexus epithelial dysfunction, illustrating the breadth and depth of the influence of KCNE2, mutations which are also associated with human cardiac arrhythmias. Here, the modus operandi and physiological roles of this potent regulator of membrane excitability and ion secretion are reviewed with particular emphasis on the ability of KCNE2 to shape the electrophysiological landscape of both excitable and non-excitable cells.
Keywords: KCNQ1; MiRP1; cardiac arrhythmia; choroid plexus; gastric acid; hypothyroidism; thyroid.
Figures