

Neurogenic subventricular zone stem/progenitor cells are Notch1-dependent in their active but not quiescent state
- PMID: 22514327
- PMCID: PMC6703480
- DOI: 10.1523/JNEUROSCI.0455-12.2012
Neurogenic subventricular zone stem/progenitor cells are Notch1-dependent in their active but not quiescent state
Abstract
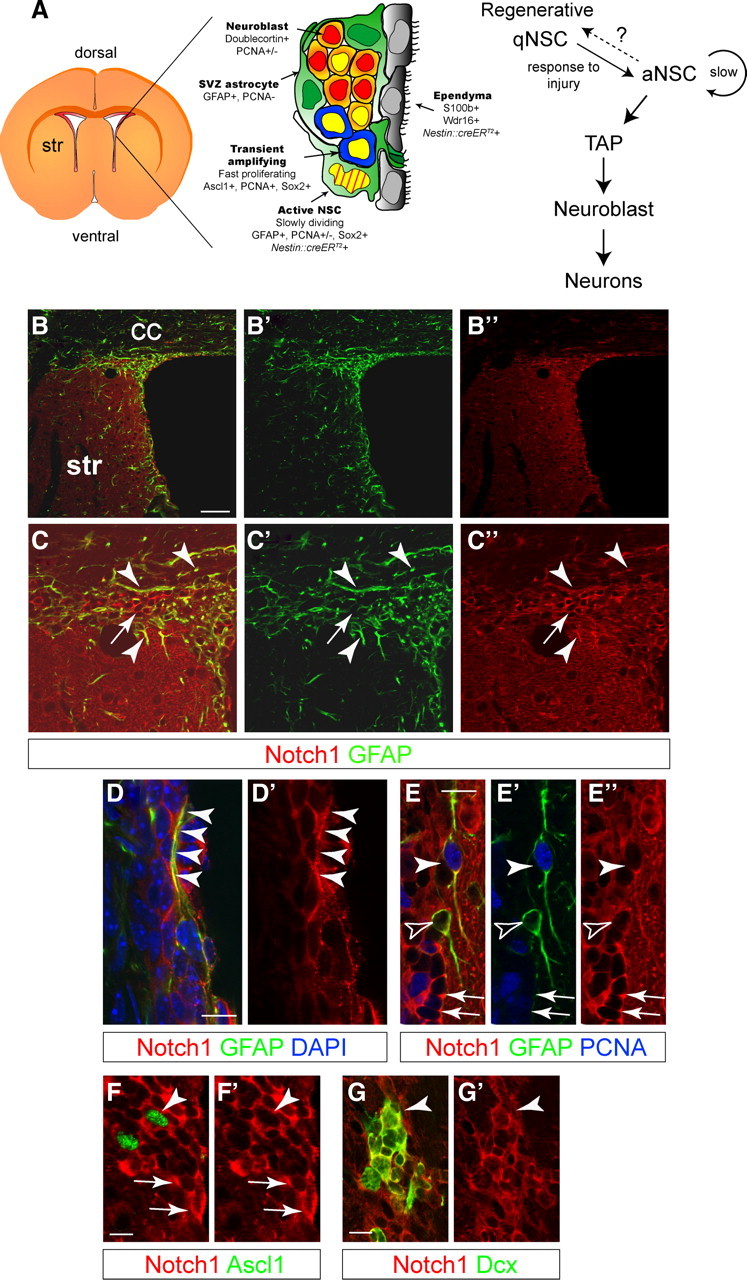
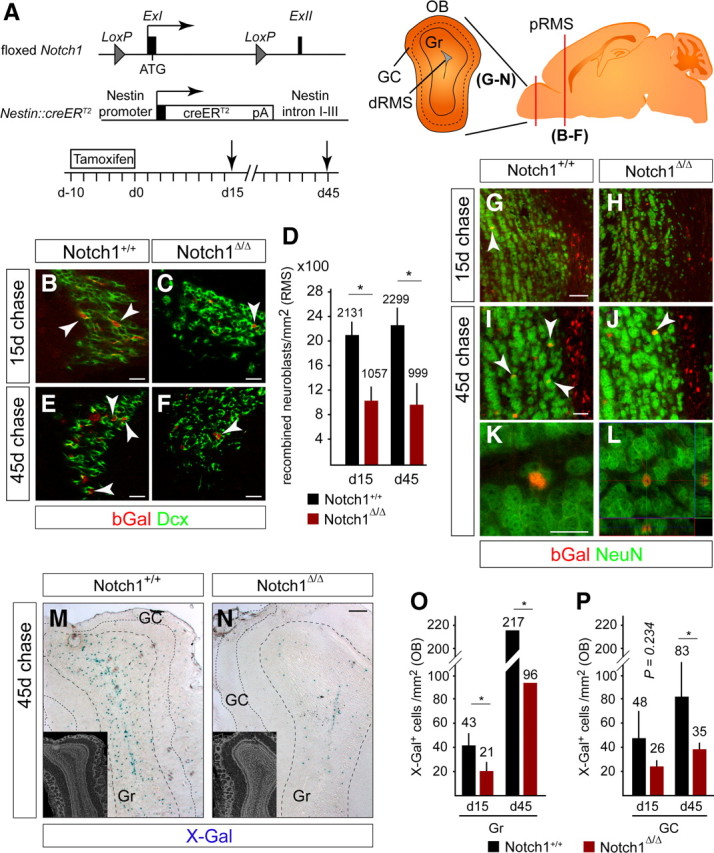
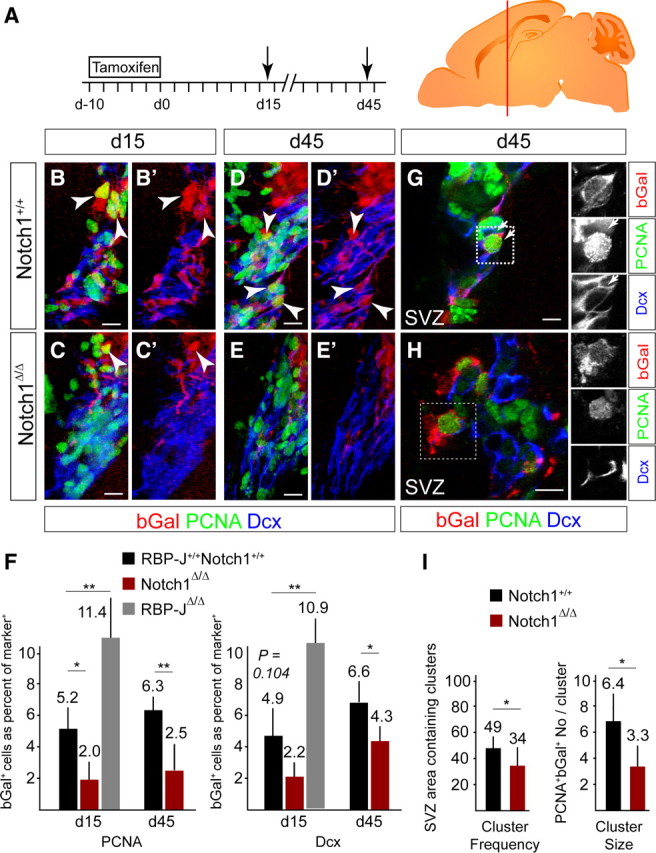
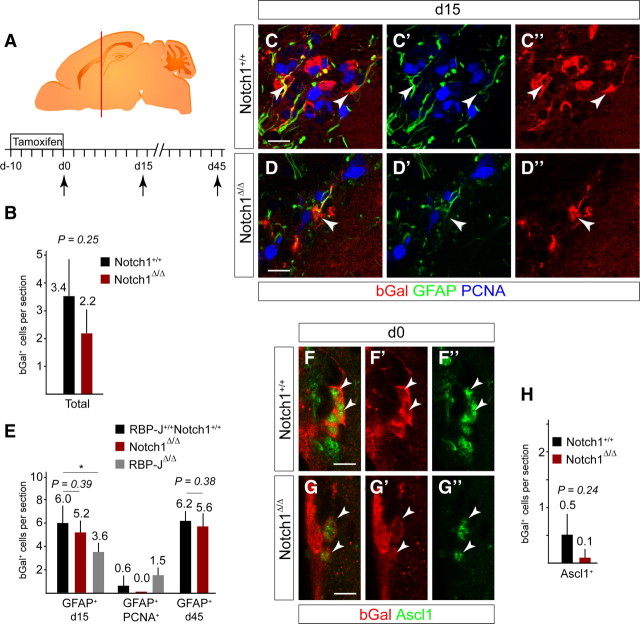
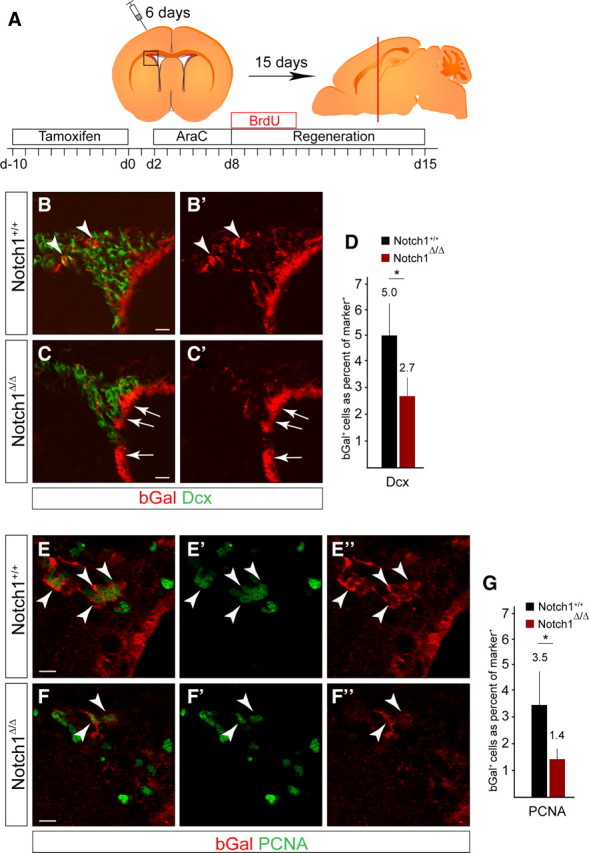
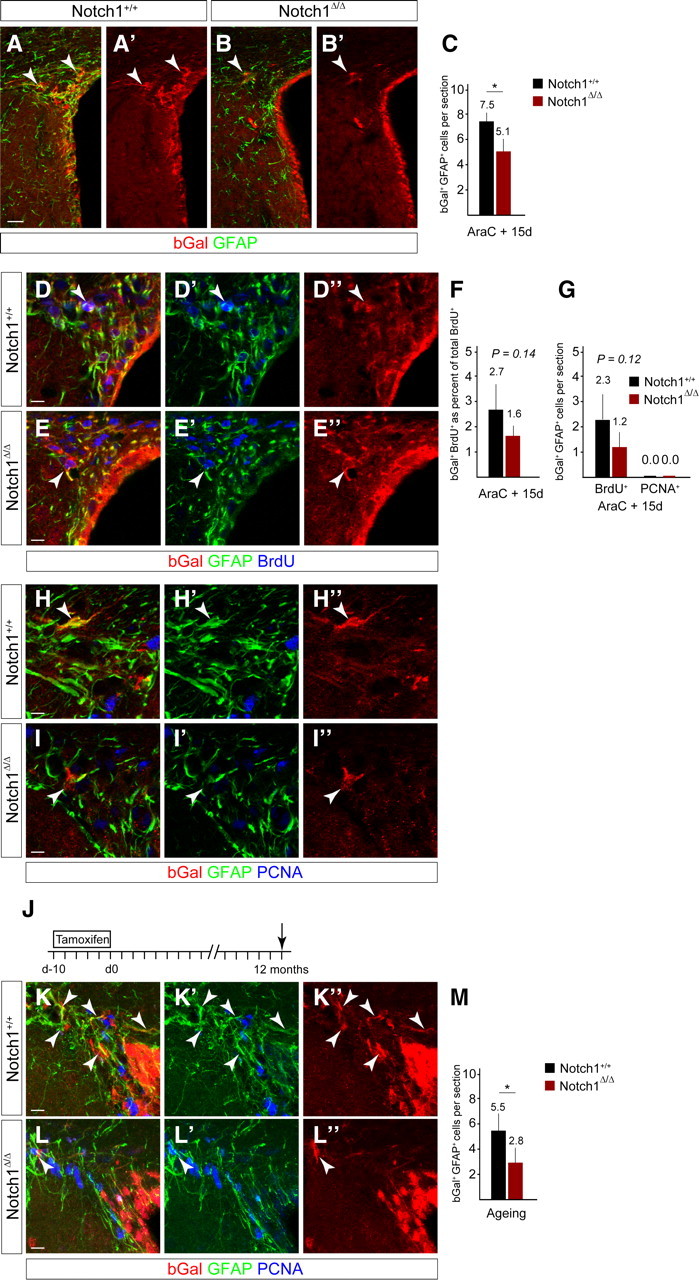
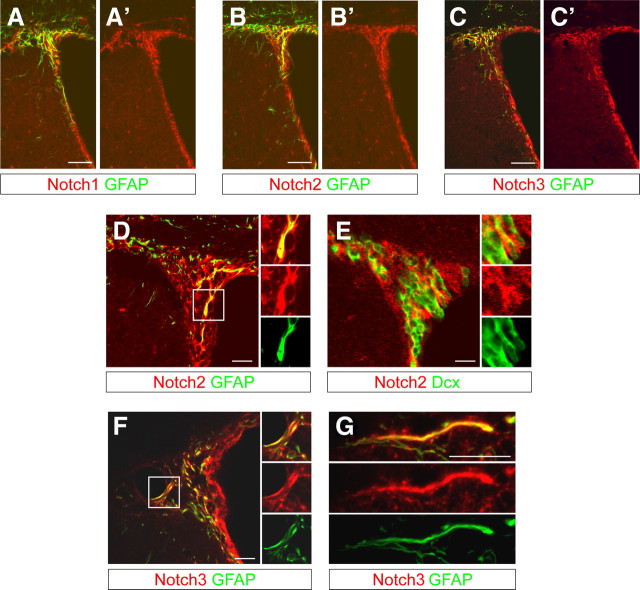
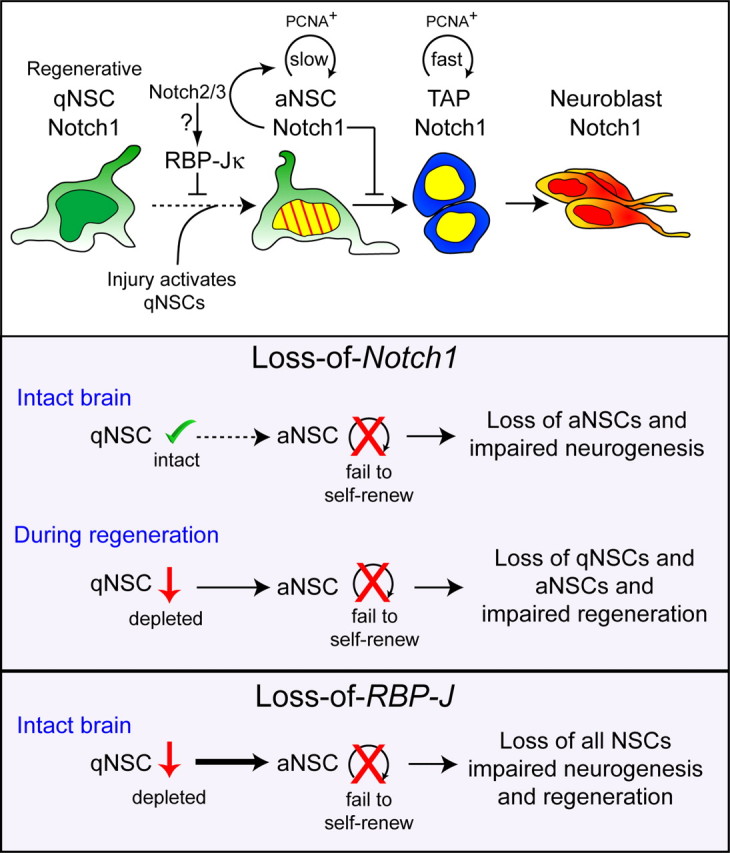
The adult mammalian forebrain contains neural stem/progenitor cells (NSCs) that generate neurons throughout life. As in other somatic stem cell systems, NSCs are proposed to be predominantly quiescent and proliferate only sporadically to produce more committed progeny. However, quiescence has recently been shown not to be an essential criterion for stem cells. It is not known whether NSCs show differences in molecular dependence based on their proliferation state. The subventricular zone (SVZ) of the adult mouse brain has a remarkable capacity for repair by activation of NSCs. The molecular interplay controlling adult NSCs during neurogenesis or regeneration is not clear but resolving these interactions is critical in order to understand brain homeostasis and repair. Using conditional genetics and fate mapping, we show that Notch signaling is essential for neurogenesis in the SVZ. By mosaic analysis, we uncovered a surprising difference in Notch dependence between active neurogenic and regenerative NSCs. While both active and regenerative NSCs depend upon canonical Notch signaling, Notch1-deletion results in a selective loss of active NSCs (aNSCs). In sharp contrast, quiescent NSCs (qNSCs) remain after Notch1 ablation until induced during regeneration or aging, whereupon they become Notch1-dependent and fail to fully reinstate neurogenesis. Our results suggest that Notch1 is a key component of the adult SVZ niche, promoting maintenance of aNSCs, and that this function is compensated in qNSCs. Therefore, we confirm the importance of Notch signaling for maintaining NSCs and neurogenesis in the adult SVZ and reveal that NSCs display a selective reliance on Notch1 that may be dictated by mitotic state.
Figures
References
-
- Basak O, Taylor V. Identification of self-replicating multipotent progenitors in the embryonic nervous system by high Notch activity and Hes5 expression. Eur J Neurosci. 2007;25:1006–1022. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials