

Acclimation of the Global Transcriptome of the Cyanobacterium Synechococcus sp. Strain PCC 7002 to Nutrient Limitations and Different Nitrogen Sources
- PMID: 22514553
- PMCID: PMC3323872
- DOI: 10.3389/fmicb.2012.00145
Acclimation of the Global Transcriptome of the Cyanobacterium Synechococcus sp. Strain PCC 7002 to Nutrient Limitations and Different Nitrogen Sources
Abstract
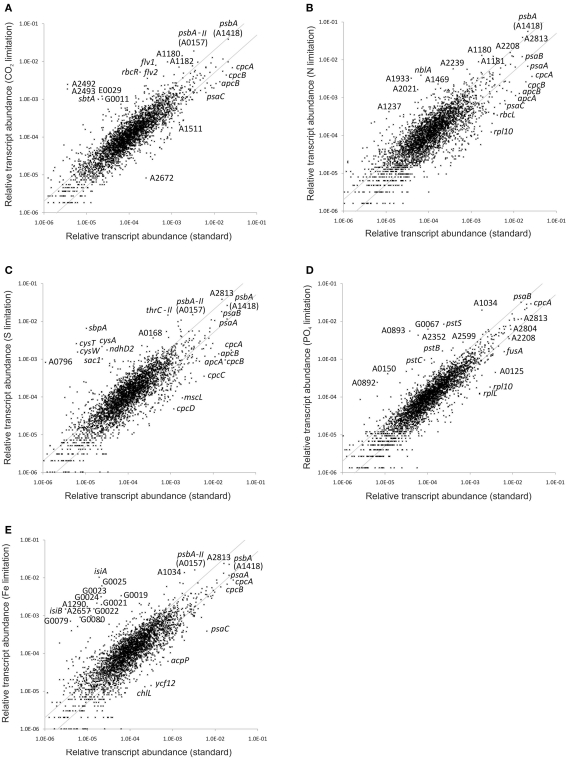
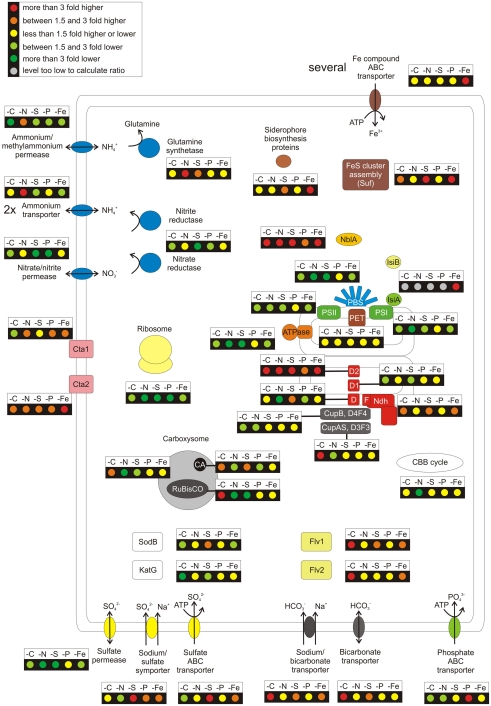
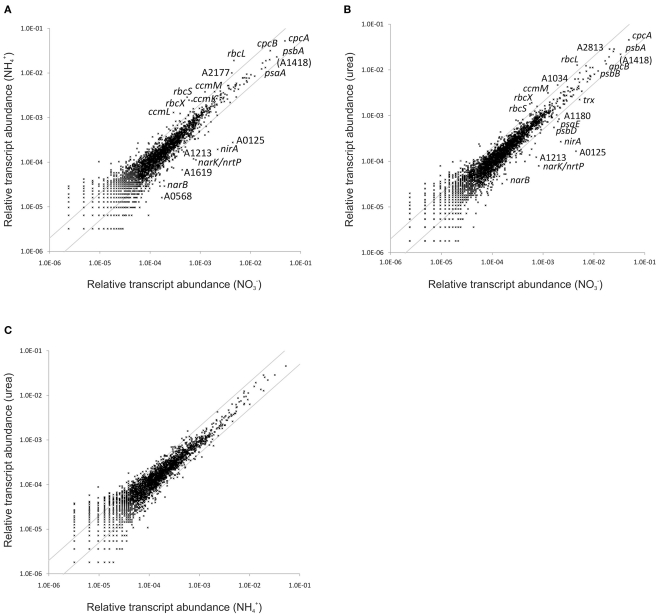
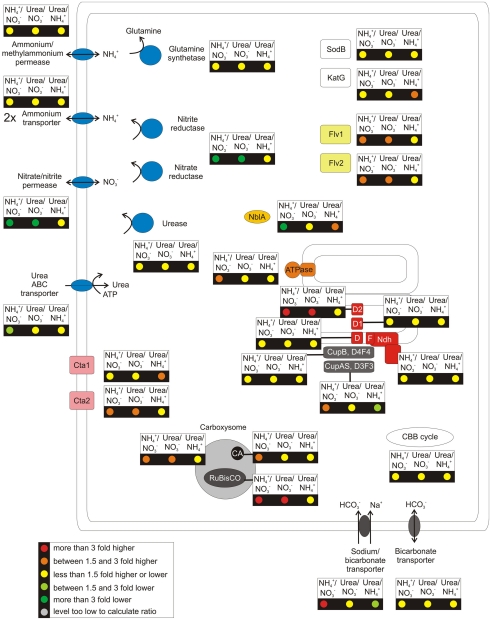
The unicellular, euryhaline cyanobacterium Synechococcus sp. strain PCC 7002 is a model organism for laboratory-based studies of cyanobacterial metabolism and is a potential platform for biotechnological applications. Two of its most notable properties are its exceptional tolerance of high-light intensity and very rapid growth under optimal conditions. In this study, transcription profiling by RNAseq has been used to perform an integrated study of global changes in transcript levels in cells subjected to limitation for the major nutrients CO(2), nitrogen, sulfate, phosphate, and iron. Transcriptional patterns for cells grown on nitrate, ammonia, and urea were also studied. Nutrient limitation caused strong decreases of transcript levels of the genes encoding major metabolic pathways, especially for components of the photosynthetic apparatus, CO(2) fixation, and protein biosynthesis. Uptake mechanisms for the respective nutrients were strongly up-regulated. The transcription data further suggest that major changes in the composition of the NADH dehydrogenase complex occur upon nutrient limitation. Transcripts for flavoproteins increased strongly when CO(2) was limiting. Genes involved in protection from oxidative stress generally showed high, constitutive transcript levels, which possibly explains the high-light tolerance of this organism. The transcriptomes of cells grown with ammonia or urea as nitrogen source showed increased transcript levels for components of the CO(2) fixation machinery compared to cells grown with nitrate, but in general transcription differences in cells grown on different N-sources exhibited surprisingly minor differences.
Keywords: RNAseq; cyanobacteria; nitrogen source; nutrient limitation; photosynthesis; transcription profiling.
Figures
References
LinkOut - more resources
Full Text Sources