

The clathrin adaptor AP-1A mediates basolateral polarity
- PMID: 22516199
- PMCID: PMC3690600
- DOI: 10.1016/j.devcel.2012.02.004
The clathrin adaptor AP-1A mediates basolateral polarity
Abstract
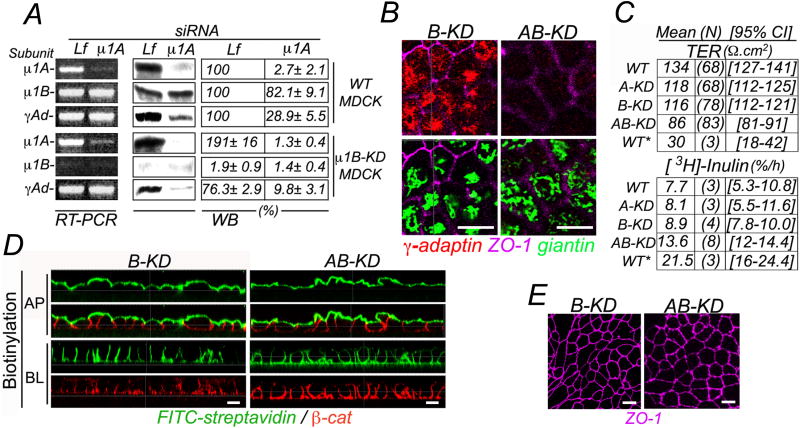
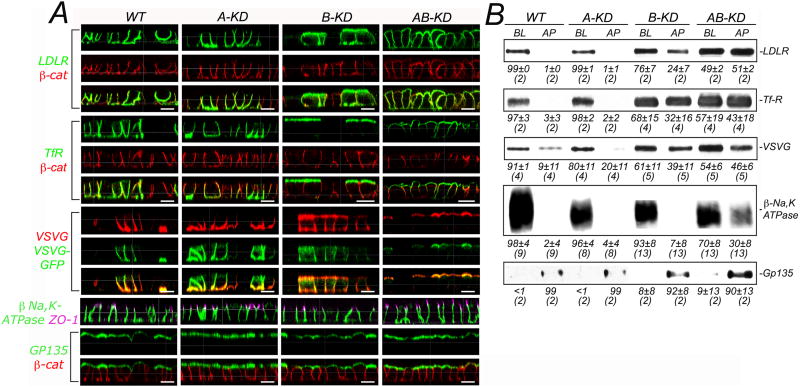
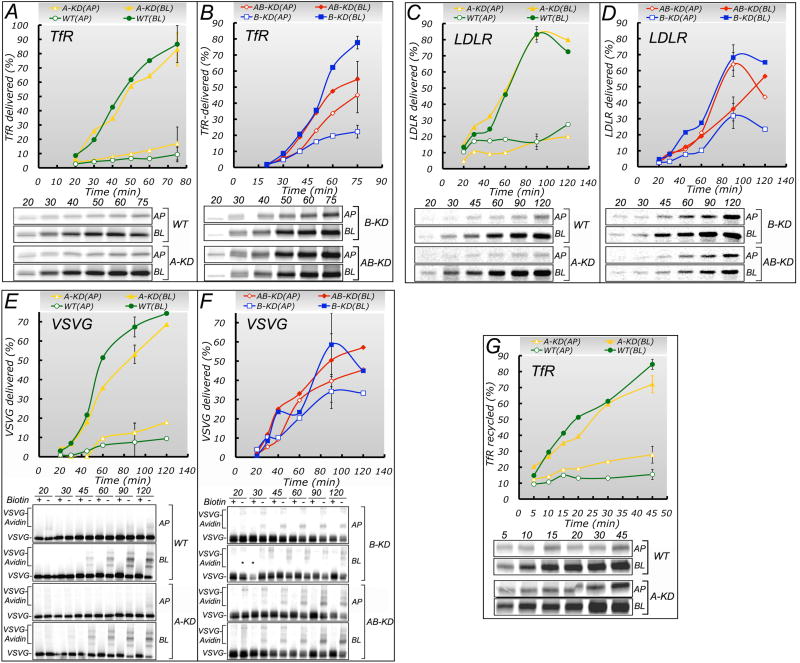
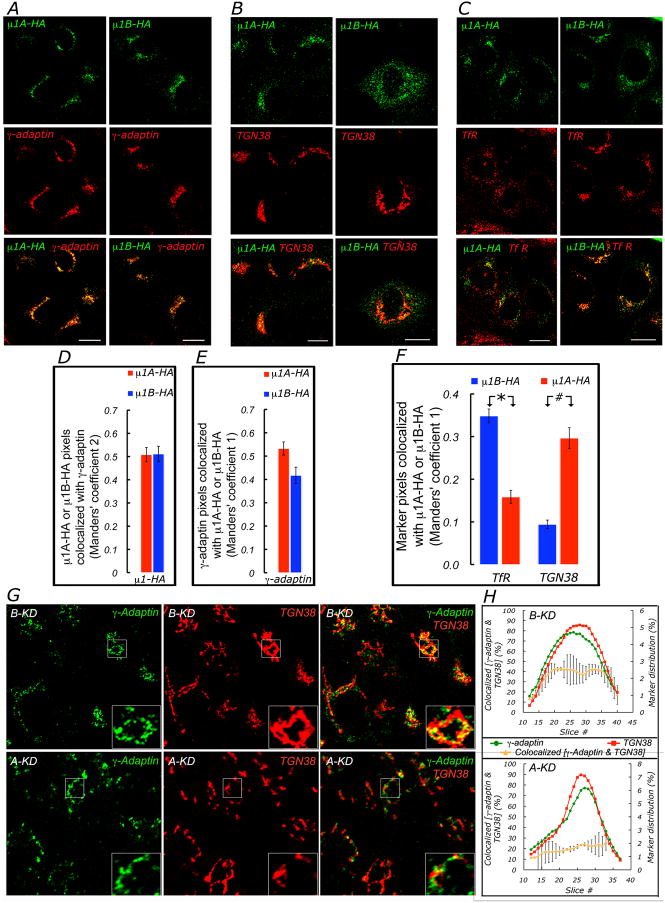
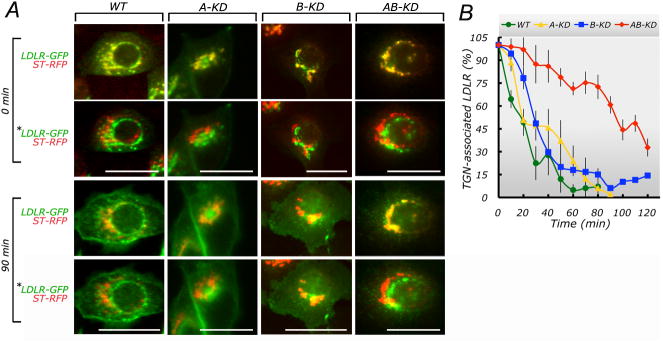
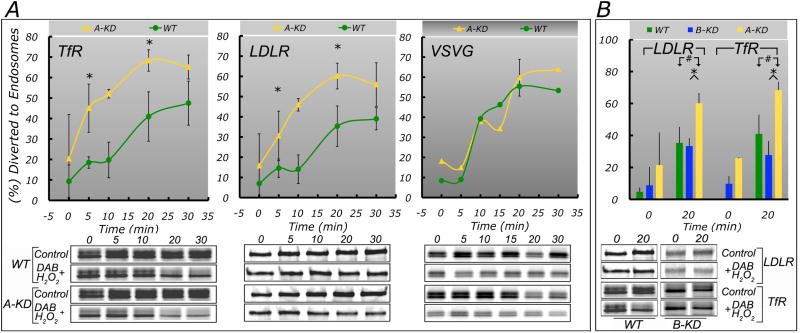
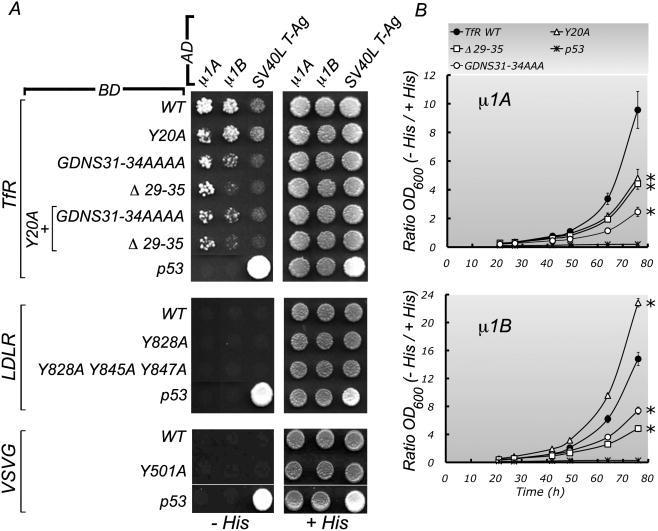
Clathrin and the epithelial-specific clathrin adaptor AP-1B mediate basolateral trafficking in epithelia. However, several epithelia lack AP-1B, and mice knocked out for AP-1B are viable, suggesting the existence of additional mechanisms that control basolateral polarity. Here, we demonstrate a distinct role of the ubiquitous clathrin adaptor AP-1A in basolateral protein sorting. Knockdown of AP-1A causes missorting of basolateral proteins in MDCK cells, but only after knockdown of AP-1B, suggesting that AP-1B can compensate for lack of AP-1A. AP-1A localizes predominantly to the TGN, and its knockdown promotes spillover of basolateral proteins into common recycling endosomes, the site of function of AP-1B, suggesting complementary roles of both adaptors in basolateral sorting. Yeast two-hybrid assays detect interactions between the basolateral signal of transferrin receptor and the medium subunits of both AP-1A and AP-1B. The basolateral sorting function of AP-1A reported here establishes AP-1 as a major regulator of epithelial polarity.
Copyright © 2012 Elsevier Inc. All rights reserved.
Figures
References
-
- Alberts B. Molecular biology of the cell. 4th. New York: Garland Science; 2002.
-
- Bonifacino JS, Traub LM. Signals for sorting of transmembrane proteins to endosomes and lysosomes. Annu Rev Biochem. 2003;72:395–447. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
