

Structural and functional characterization of microcin C resistance peptidase MccF from Bacillus anthracis
- PMID: 22516613
- PMCID: PMC3690760
- DOI: 10.1016/j.jmb.2012.04.011
Structural and functional characterization of microcin C resistance peptidase MccF from Bacillus anthracis
Abstract
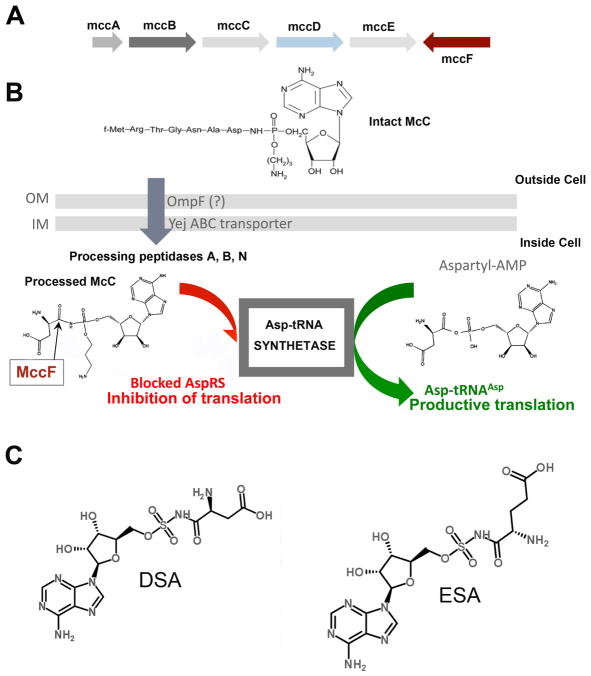
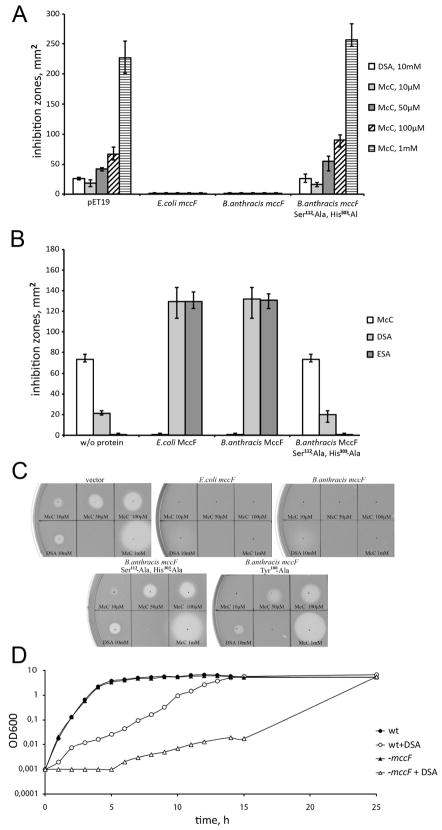
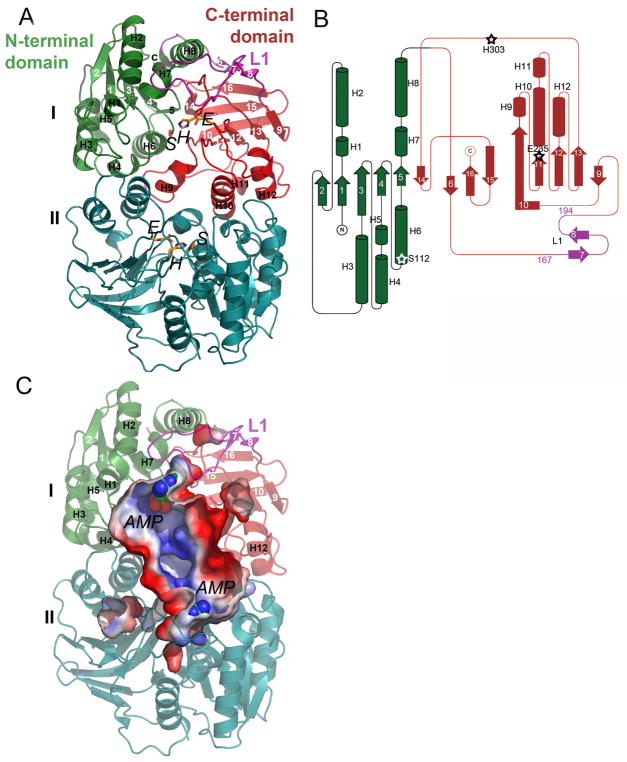
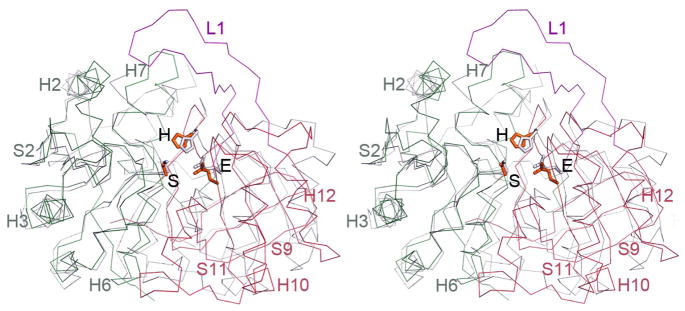
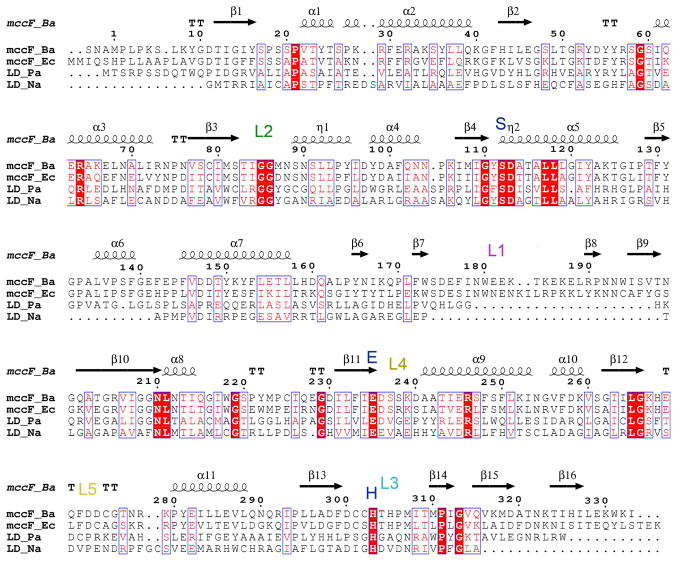
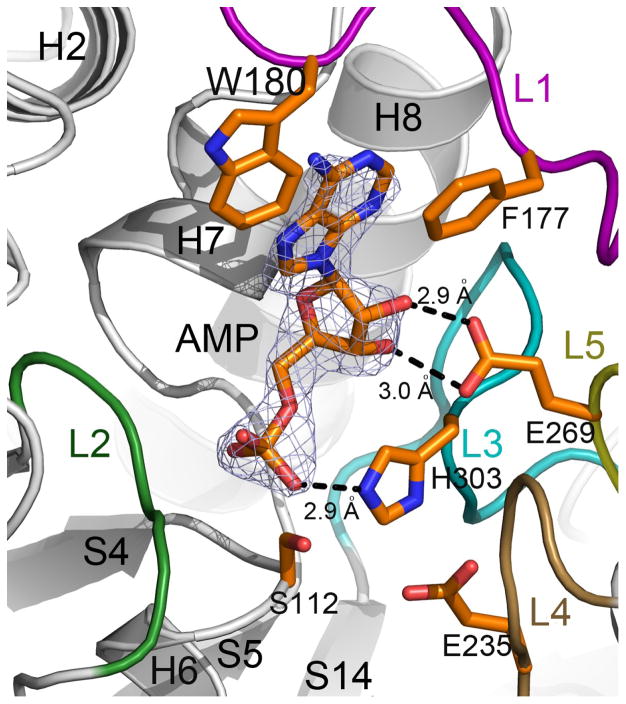
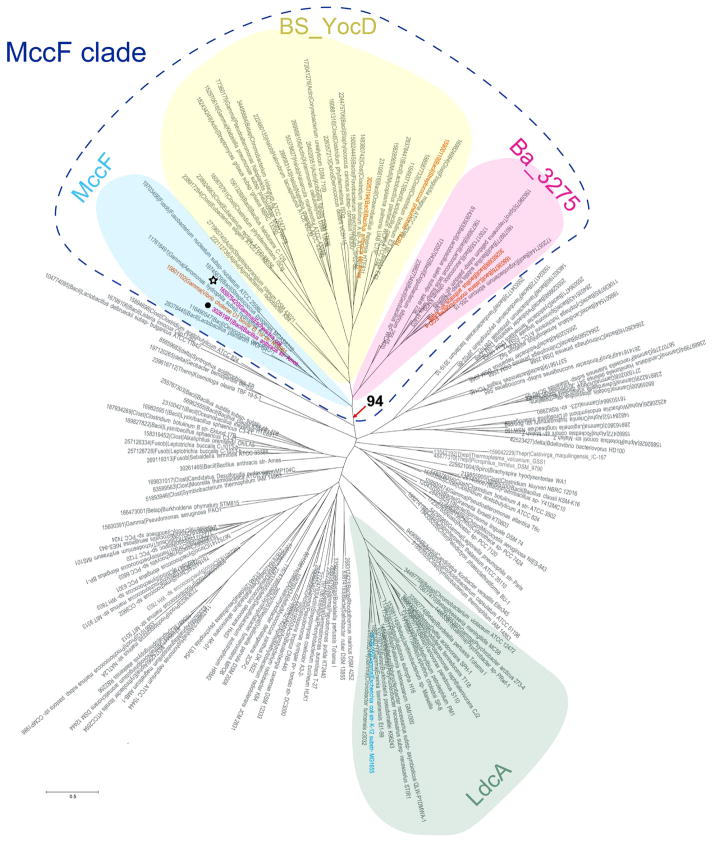
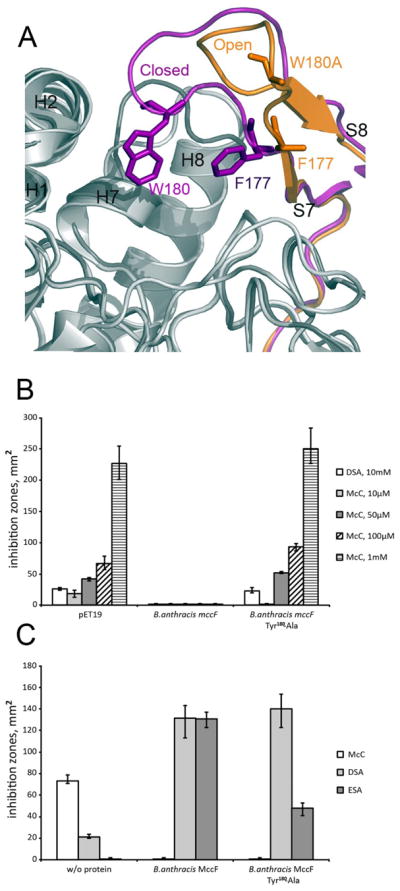
Microcin C (McC) is heptapeptide adenylate antibiotic produced by Escherichia coli strains carrying the mccABCDEF gene cluster encoding enzymes, in addition to the heptapeptide structural gene mccA, necessary for McC biosynthesis and self-immunity of the producing cell. The heptapeptide facilitates McC transport into susceptible cells, where it is processed releasing a non-hydrolyzable aminoacyl adenylate that inhibits an essential aminoacyl-tRNA synthetase. The self-immunity gene mccF encodes a specialized serine peptidase that cleaves an amide bond connecting the peptidyl or aminoacyl moieties of, respectively, intact and processed McC with the nucleotidyl moiety. Most mccF orthologs from organisms other than E. coli are not linked to the McC biosynthesis gene cluster. Here, we show that a protein product of one such gene, MccF from Bacillus anthracis (BaMccF), is able to cleave intact and processed McC, and we present a series of structures of this protein. Structural analysis of apo-BaMccF and its adenosine monophosphate complex reveals specific features of MccF-like peptidases that allow them to interact with substrates containing nucleotidyl moieties. Sequence analyses and phylogenetic reconstructions suggest that several distinct subfamilies form the MccF clade of the large S66 family of bacterial serine peptidases. We show that various representatives of the MccF clade can specifically detoxify non-hydrolyzable aminoacyl adenylates differing in their aminoacyl moieties. We hypothesize that bacterial mccF genes serve as a source of bacterial antibiotic resistance.
Copyright © 2012 Elsevier Ltd. All rights reserved.
Figures
References
-
- Severinov K, Semenova E, Kazakov A, Kazakov T, Gelfand MS. Low-molecular-weight post-translationally modified microcins. Mol Microbiol. 2007;65:1380–94. - PubMed
-
- Destoumieux-Garzon D, Peduzzi J, Rebuffat S. Focus on modified microcins: structural features and mechanisms of action. Biochimie. 2002;84:511–9. - PubMed
-
- Guijarro JI, Gonzalez-Pastor JE, Baleux F, San Millan JL, Castilla MA, Rico M, Moreno F, Delepierre M. Chemical structure and translation inhibition studies of the antibiotic microcin C7. J Biol Chem. 1995;270:23520–32. - PubMed
-
- Metlitskaya AZ, Katrukha GS, Shashkov AS, Zaitsev DA, Egorov TA, Khmel IA. Structure of microcin C51, a new antibiotic with a broad spectrum of activity. FEBS Lett. 1995;357:235–8. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
