

Regulation and function of mTOR signalling in T cell fate decisions
- PMID: 22517423
- PMCID: PMC3417069
- DOI: 10.1038/nri3198
Regulation and function of mTOR signalling in T cell fate decisions
Abstract
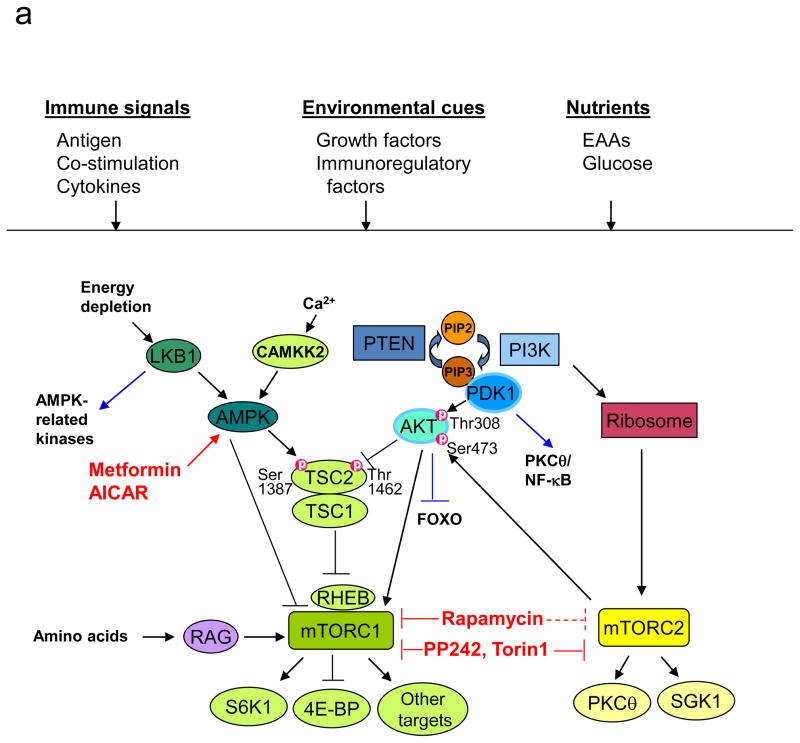
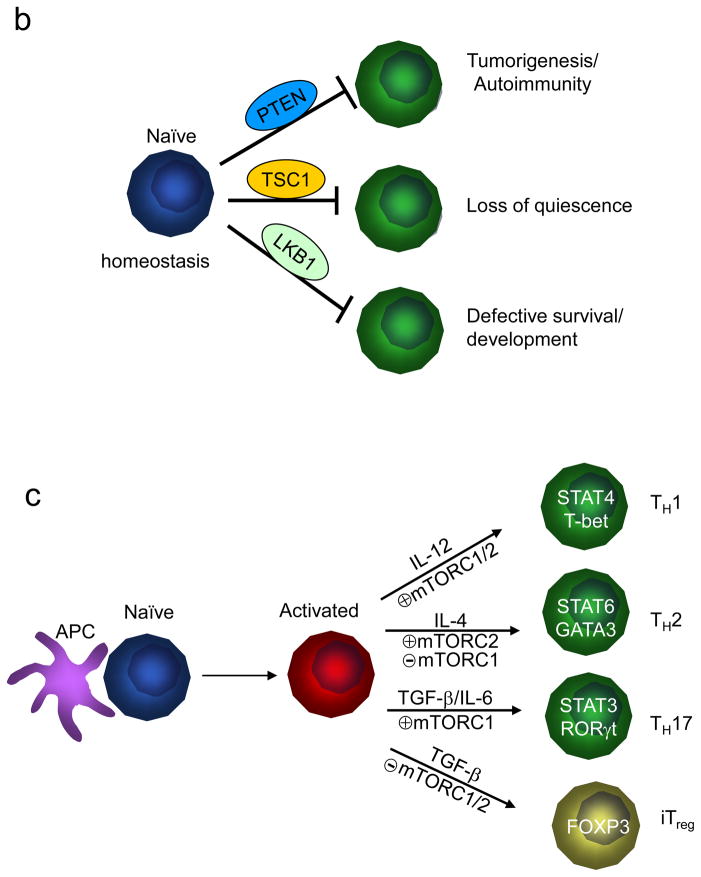
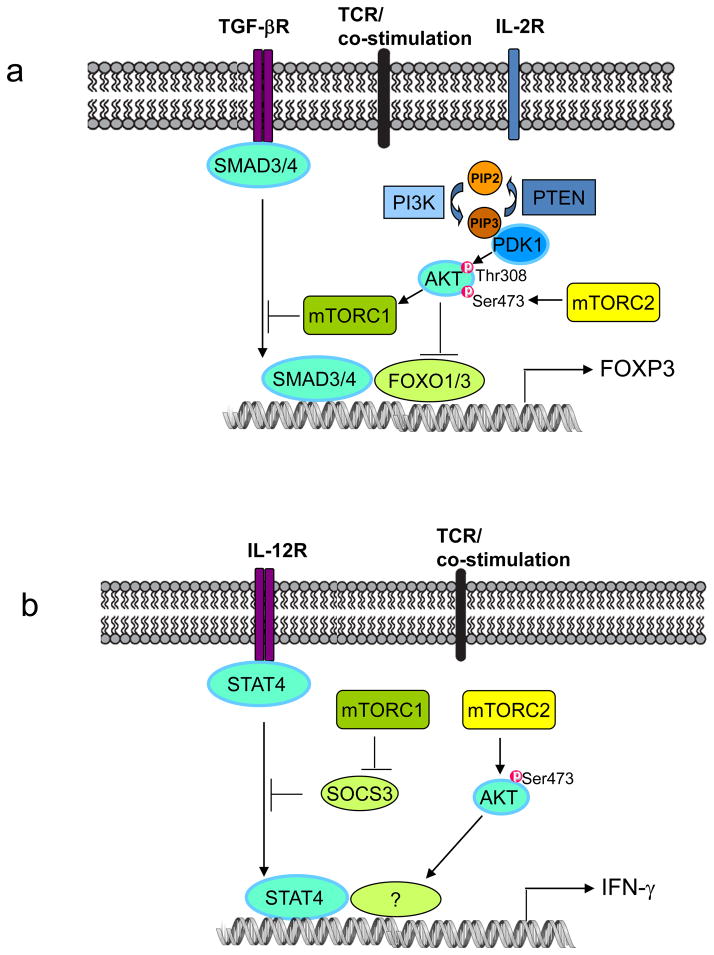
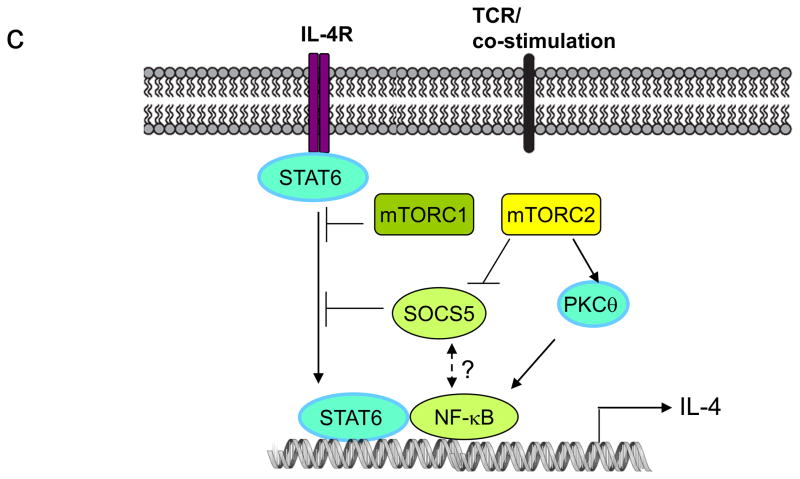
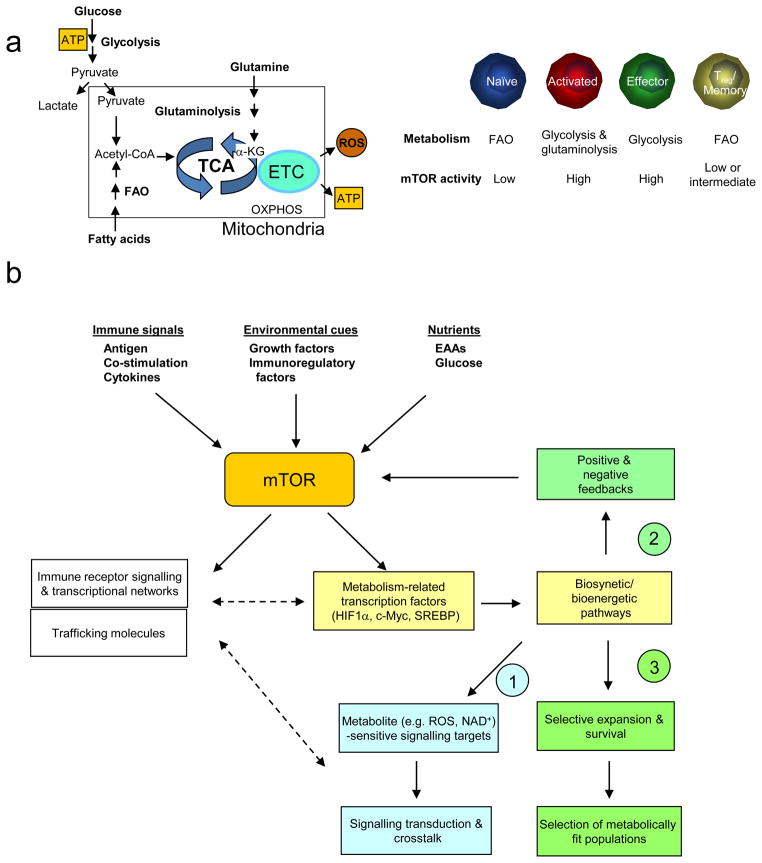
The evolutionarily conserved kinase mTOR (mammalian target of rapamycin) couples cell growth and metabolism to environmental inputs in eukaryotes. T cells depend on mTOR signalling to integrate immune signals and metabolic cues for their proper maintenance and activation. Under steady-state conditions, mTOR is actively controlled by multiple inhibitory mechanisms, and this enforces normal T cell homeostasis. Antigen recognition by naive CD4(+) and CD8(+) T cells triggers mTOR activation, which in turn programmes the differentiation of these cells into functionally distinct lineages. This Review focuses on the signalling mechanisms of mTOR in T cell homeostatic and functional fates, and discusses the therapeutic implications of targeting mTOR in T cells.
Conflict of interest statement
The author declares no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
