

Water status and associated processes mark critical stages in pollen development and functioning
- PMID: 22523424
- PMCID: PMC3359924
- DOI: 10.1093/aob/mcs070
Water status and associated processes mark critical stages in pollen development and functioning
Abstract
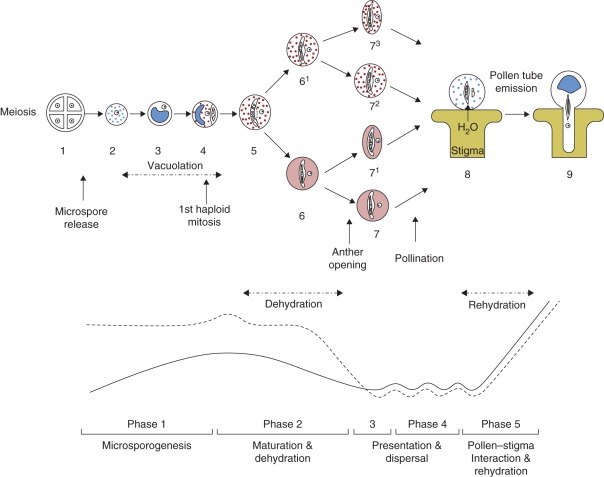
Background: The male gametophyte developmental programme can be divided into five phases which differ in relation to the environment and pollen hydration state: (1) pollen develops inside the anther immersed in locular fluid, which conveys substances from the mother plant--the microsporogenesis phase; (2) locular fluid disappears by reabsorption and/or evaporation before the anther opens and the maturing pollen grains undergo dehydration--the dehydration phase; (3) the anther opens and pollen may be dispersed immediately, or be held by, for example, pollenkitt (as occurs in almost all entomophilous species) for later dispersion--the presentation phase; (4) pollen is dispersed by different agents, remaining exposed to the environment for different periods--the dispersal phase; and (5) pollen lands on a stigma and, in the case of a compatible stigma and suitable conditions, undergoes rehydration and starts germination--the pollen-stigma interaction phase.
Scope: This review highlights the issue of pollen water status and indicates the various mechanisms used by pollen grains during their five developmental phases to adjust to changes in water content and maintain internal stability.
Conclusions: Pollen water status is co-ordinated through structural, physiological and molecular mechanisms. The structural components participating in regulation of the pollen water level, during both dehydration and rehydration, include the exine (the outer wall of the pollen grain) and the vacuole. Recent data suggest the involvement of water channels in pollen water transport and the existence of several molecular mechanisms for pollen osmoregulation and to protect cellular components (proteins and membranes) under water stress. It is suggested that pollen grains will use these mechanisms, which have a developmental role, to cope with environmental stress conditions.
Figures


References
-
- Aarts MG, Hodge R, Kalantidis K, et al. The Arabidopsis MALE STERILITY 2 protein shares similarity with reductases in elongation/condensation complexes. The Plant Journal. 1997;12:615–623. - PubMed
-
- Abba S, Ghignone S, Bonfante P. A dehydration-inducible gene in the truffle Tuber borchii identifies a novel group of dehydrins. BMC Genomics. 2006;7(39) http://dx.doi.org/10.1186/1471-2164-7-39 . - PMC - PubMed
-
- Agarwal PK, Agarwal P, Reddy MK, Sopory SK. Role of DREB transcription factors in abiotic and biotic stress tolerance in plants. Plant Cell Reports. 2006;25:1263–1274. - PubMed
-
- Alpert P, Oliver MJ. Drying without dying. In: Black M, Pritchard HW, editors. Dessication and survival in plants. Wallingford, UK: CABI Publishing; 2002. pp. 3–43.
-
- Ariizumi T, Hatakeyama K, Hinata K, et al. A novel male-sterile mutant of Arabidopsis thaliana, faceless pollen-1, produces pollen with a smooth surface and an acetolysis-sensitive exine. Plant Molecular Biology. 2003;53:107–116. - PubMed
