

Developmental regulation of chromatin conformation by Hox proteins in Drosophila
- PMID: 22523743
- PMCID: PMC3329935
- DOI: 10.1016/j.celrep.2012.03.003
Developmental regulation of chromatin conformation by Hox proteins in Drosophila
Abstract
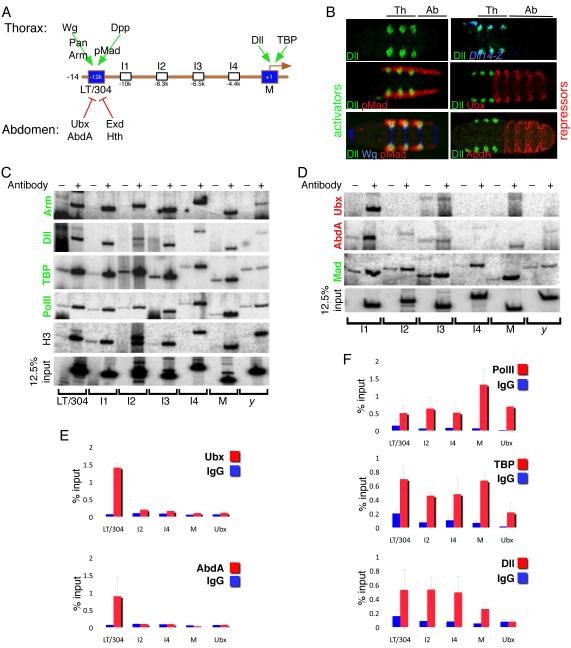
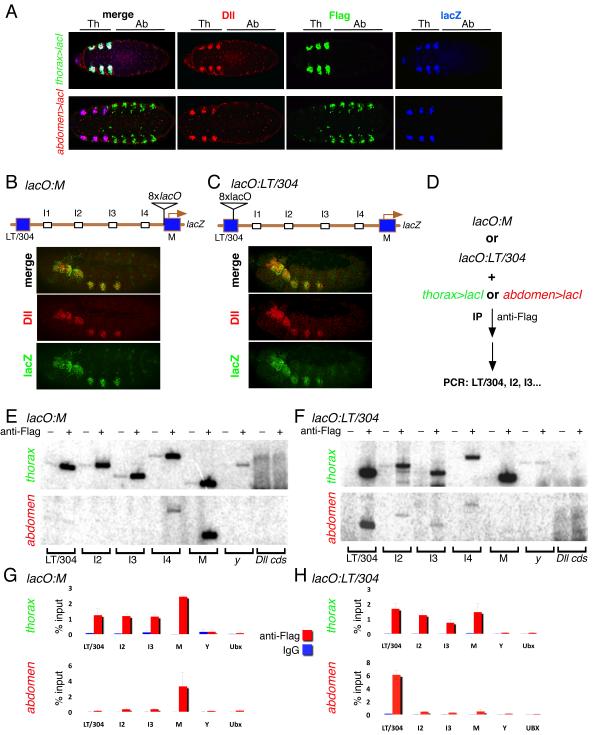
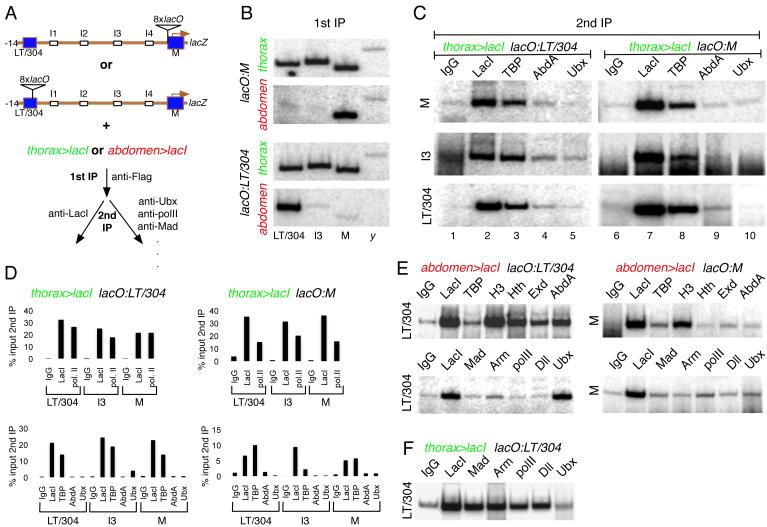
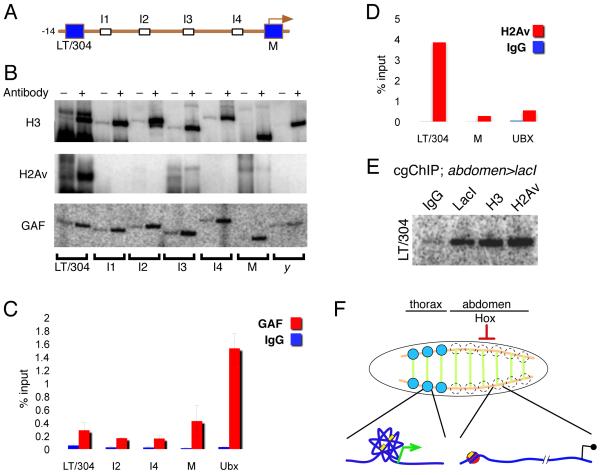
We present a strategy to examine the chromatin conformation of individual loci in specific cell types during Drosophila embryogenesis. Regulatory DNA is tagged with binding sites (lacO) for LacI, which is used to immunoprecipitate the tagged chromatin from specific cell types. We applied this approach to Distalless (Dll), a gene required for limb development in Drosophila. We show that the local chromatin conformation at Dll depends on the cell type: in cells that express Dll, the 5' regulatory region is in close proximity to the Dll promoter. In Dll-nonexpressing cells this DNA is in a more extended configuration. In addition, transcriptional activators and repressors are bound to Dll regulatory DNA in a cell type-specific manner. The pattern of binding by GAGA factor and the variant histone H2Av suggest that they play a role in the regulation of Dll chromatin conformation in expressing and nonexpressing cell types, respectively.
Figures
References
-
- Amano T, Sagai T, Tanabe H, Mizushina Y, Nakazawa H, Shiroishi T. Chromosomal dynamics at the Shh locus: limb bud-specific differential regulation of competence and active transcription. Dev Cell. 2009;16:47–57. - PubMed
-
- Apostolou E, Thanos D. Virus Infection Induces NF-kappaB-dependent interchromosomal associations mediating monoallelic IFN-beta gene expression. Cell. 2008;134:85–96. - PubMed
-
- Bartkuhn M, Renkawitz R. Long range chromatin interactions involved in gene regulation. Biochim Biophys Acta. 2008;1783:2161–2166. - PubMed
-
- Biggin MD. Animal transcription networks as highly connected, quantitative continua. Dev Cell. 2011;21:611–626. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
