

Redox-sensitive GFP2: use of the genetically encoded biosensor of the redox status in the filamentous fungus Botrytis cinerea
- PMID: 22524254
- PMCID: PMC6638776
- DOI: 10.1111/j.1364-3703.2012.00802.x
Redox-sensitive GFP2: use of the genetically encoded biosensor of the redox status in the filamentous fungus Botrytis cinerea
Abstract
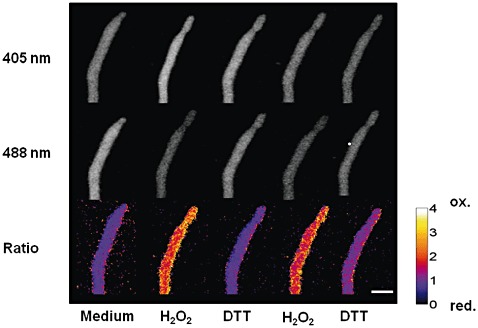
The production of reactive oxygen species (ROS) is part of the defence reaction of plants against invading pathogens. The effect of ROS on filamentous fungi is still unclear. In this study, ratiometric redox-sensitive green fluorescent protein (roGFP) was introduced as a tool for in vivo measurement of the cellular redox status in filamentous fungi. A fungal expression system for roGFP2 was constructed. Expressed in Botrytis cinerea, roGFP2 reversibly responded to redox changes induced by incubation with H(2)O(2) or dithiothreitol, which was determined by confocal laser scanning microscopy imaging and fluorometry. As the sensor detects the redox potential of the cellular glutathione pool, it was used to analyse the kinetics of GSH (glutathione, reduced form) recovery after H(2)O(2) treatment. The transcription factor Bap1 is the main transcriptional regulator of H(2)O(2) -scavenging proteins in B. cinerea. When compared with the wild-type, GSH recovery in the Δbap1 deletion mutant was affected after repeated H(2)O(2) treatment. ROS and intracellular redox changes can be used by fungi for signalling purposes. In planta experiments, performed in this study, indicated that redox processes seem to be important for the differentiation of penetration structures. During the penetration of onion epidermal cells, the status of the cellular glutathione pool differed between appressoria-like structures and infecting hyphae, being reduced in the presence of infecting hyphae and more oxidized around appressoria-like structures.
© 2012 THE AUTHORS. MOLECULAR PLANT PATHOLOGY © 2012 BSPP AND BLACKWELL PUBLISHING LTD.
Figures







References
-
- Aguirre, J. , Ríos‐Momberg, M. , Hewitt, D. and Hansberg, W. (2005) Reactive oxygen species and development in microbial eukaryotes. Trends Microbiol. 13, 111–118. - PubMed
-
- Brun, S. , Malagnac, F. , Bidard, F. , Lalucque, H. and Silar, P. (2009) Functions and regulation of the nox family in the filamentous fungus Podospora anserina: a new role in cellulose degradation. Mol. Microbiol. 74, 480–496. - PubMed
-
- Büttner, P. , Koch, F. , Voigt, K. , Quidde, T. , Risch, S. , Blaich, R. , Brückner, B. and Tudzynski, P. (1994) Variations in ploidy among isolates of Botrytis cinerea: implications for genetic and molecular analyses. Curr. Genet. 25, 445–450. - PubMed
-
- Colot, H.V. , Park, G. , Turner, G.E. , Ringelberg, C. , Crew, C.M. , Litvinkova, L. , Weiss, R.L. , Borkovich, K.A. and Dunlap, J.C. (2006) A high‐throughput gene knockout procedure for Neurospora reveals functions for multiple transcription factors. Proc. Natl. Acad. Sci. USA, 103, 10 352–10 357. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
