

Interkinetic nuclear migration: a mysterious process in search of a function
- PMID: 22524603
- PMCID: PMC3357188
- DOI: 10.1111/j.1440-169X.2012.01342.x
Interkinetic nuclear migration: a mysterious process in search of a function
Abstract
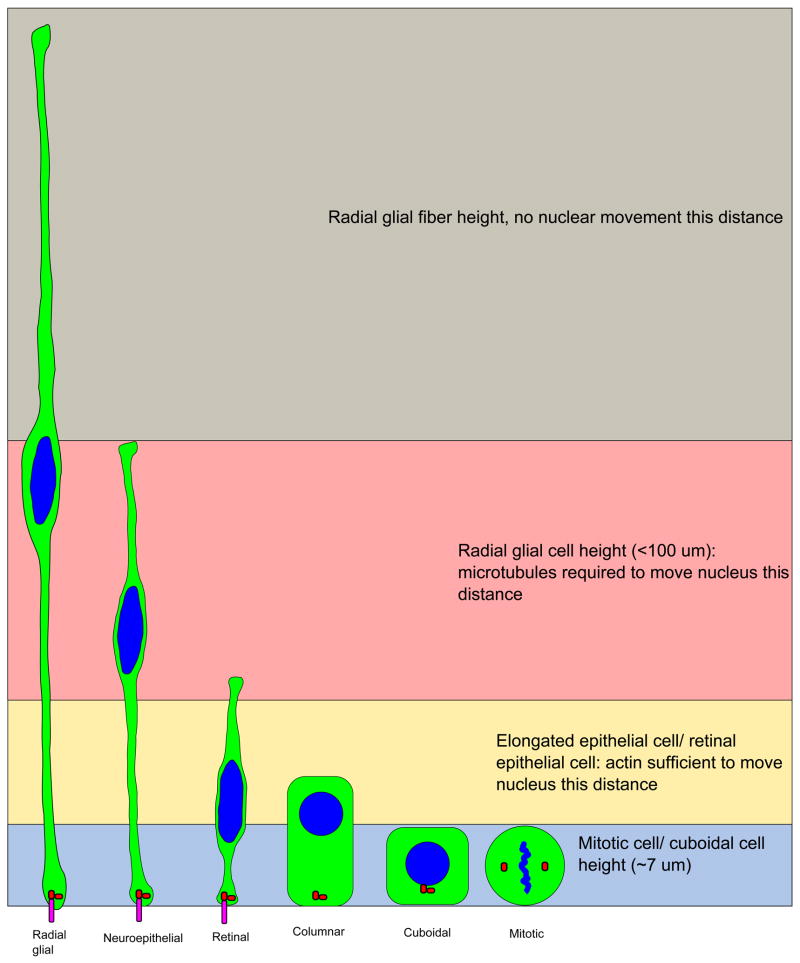
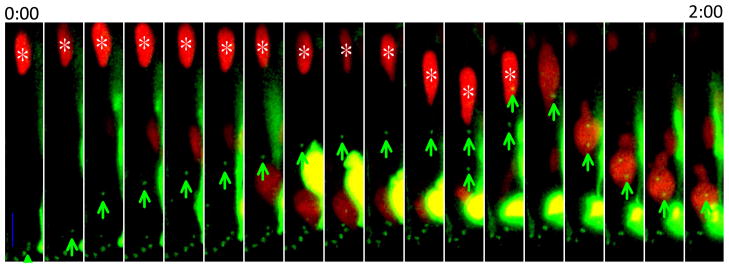
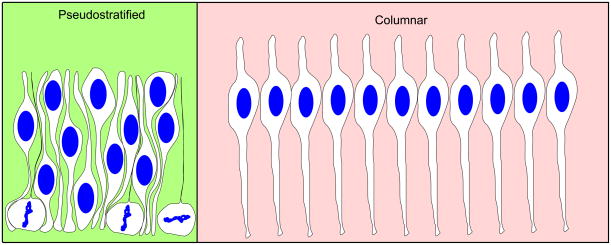
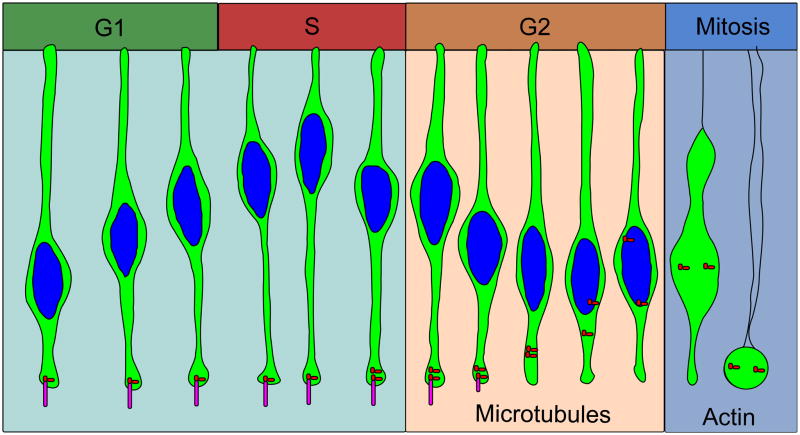
During interkinetic nuclear migration (INM), the nuclei in many epithelial cells migrate between the apical and basal surfaces, coordinating with the cell cycle, and undergoing cytokinesis at the apical surface. INM is observed in a wide variety of tissues and species. Recent advances in time-lapse microscopy have provided clues about the mechanisms and functions of INM. Whether actin or microtubules are responsible for nuclear migration is controversial. How mitosis is initiated during INM is poorly understood, as is the relationship between the cell cycle and nuclear movement. It is possible that the disagreements stem from differences in the tissues being studied, since epithelia undergoing INM vary greatly in terms of cell height and cell fates. In this review we examine the reports addressing the mode and mechanisms that regulate INM and suggest possible functions for this dramatic event.
Published 2012. This article is a US Government work and is in the public domain in the USA Development, Growth & Differentiation © 2012 Japanese Society of Developmental Biologists.
Figures
References
-
- Afonso C, Henrique D. PAR3 acts as a molecular organizer to define the apical domain of chick neuroepithelial cells. J Cell Sci. 2006;119:4293–304. - PubMed
-
- Austin CP, Feldman DE, Ida JA, Jr, Cepko CL. Vertebrate retinal ganglion cells are selected from competent progenitors by the action of Notch. Development. 1995;121:3637–3650. - PubMed
-
- Basto R, Pines J. The Centrosome Opens the Way to Mitosis. Developmental Cell. 2007;12:475–477. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
