

Induction of Bv8 expression by granulocyte colony-stimulating factor in CD11b+Gr1+ cells: key role of Stat3 signaling
- PMID: 22528488
- PMCID: PMC3365993
- DOI: 10.1074/jbc.M111.326801
Induction of Bv8 expression by granulocyte colony-stimulating factor in CD11b+Gr1+ cells: key role of Stat3 signaling
Abstract
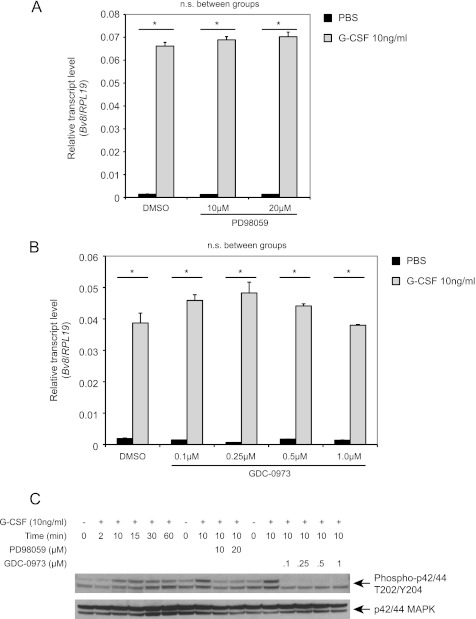
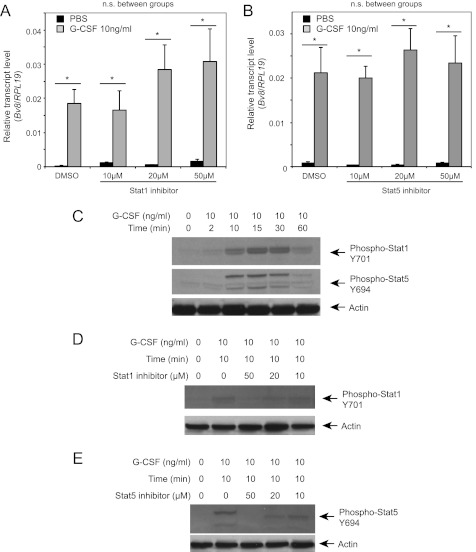
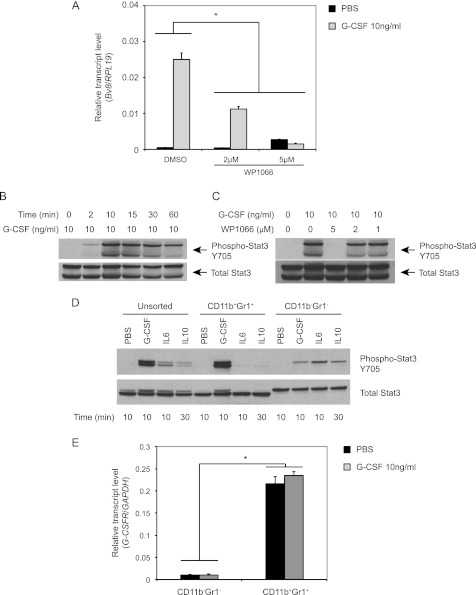
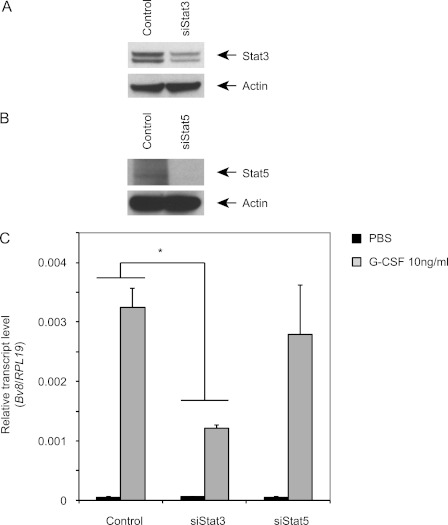
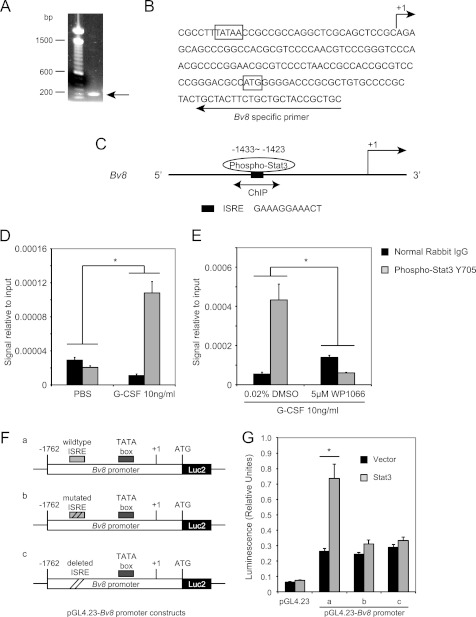
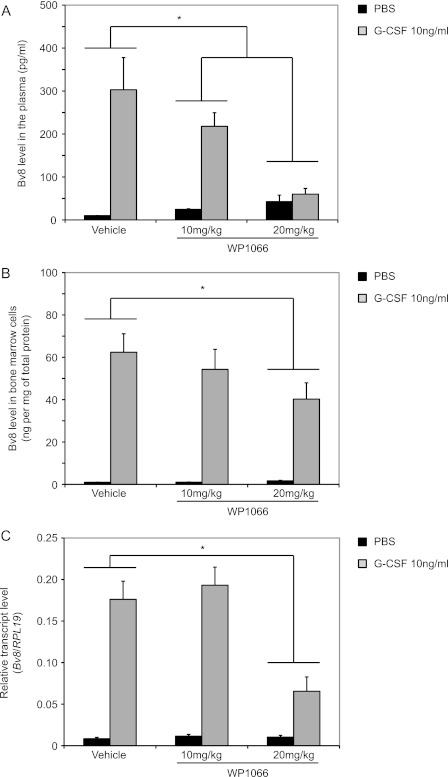
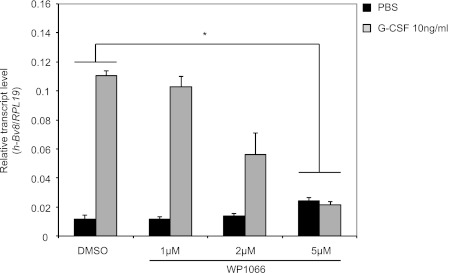
Bv8, also known as prokineticin 2, has been characterized as an important mediator of myeloid cell mobilization and myeloid cell-dependent tumor angiogenesis. Bv8 expression is dramatically enhanced by G-CSF, both in vitro and in vivo. The mechanisms involved in such up-regulation remain unknown. Using pharmacological inhibitors that interfere with multiple signaling pathways known to be activated by G-CSF, we show that signal transducer and activator of transcription 3 (Stat3) activation is required for Bv8 up-regulation in mouse bone marrow cells, whereas other Stat family members and extracellular signal-regulated kinase (ERK) activation are not involved. We further identified CD11b(+) Gr1(+) myeloid cells as the primary cell population in which Stat3 signaling is activated by G-CSF. Bv8 expression induced by G-CSF was also significantly reduced by siRNA-mediated Stat3 knockdown. Moreover, chromatin immunoprecipitation studies indicate that G-CSF significantly induces binding of phospho-Stat3 to the Bv8 promoter, which was abolished by pretreatment with the Stat3 inhibitor WP1066. Luciferase assay confirmed that the phospho-Stat3 binding site is a functional enhancer of the Bv8 promoter. The key role of Stat3 signaling in regulating G-CSF-induced Bv8 expression was further confirmed by in vivo studies. We show that the regulation of Bv8 expression in human bone marrow cells is also Stat3 signaling-dependent. Stat3 is recognized as a key regulator of inflammation-dependent tumorigenesis. We propose that such a role of Stat3 reflects at least in part its ability to regulate Bv8 expression.
Figures
References
-
- Li M., Bullock C. M., Knauer D. J., Ehlert F. J., Zhou Q. Y. (2001) Identification of two prokineticin cDNAs: recombinant proteins potently contract gastrointestinal smooth muscle. Mol. Pharmacol. 59, 692–698 - PubMed
-
- LeCouter J., Kowalski J., Foster J., Hass P., Zhang Z., Dillard-Telm L., Frantz G., Rangell L., DeGuzman L., Keller G. A., Peale F., Gurney A., Hillan K. J., Ferrara N. (2001) Identification of an angiogenic mitogen selective for endocrine gland endothelium. Nature 412, 877–884 - PubMed
-
- Mollay C., Wechselberger C., Mignogna G., Negri L., Melchiorri P., Barra D., Kreil G. (1999) Bv8, a small protein from frog skin and its homologue from snake venom induce hyperalgesia in rats. Eur. J. Pharmacol. 374, 189–196 - PubMed
-
- Masuda Y., Takatsu Y., Terao Y., Kumano S., Ishibashi Y., Suenaga M., Abe M., Fukusumi S., Watanabe T., Shintani Y., Yamada T., Hinuma S., Inatomi N., Ohtaki T., Onda H., Fujino M. (2002) Isolation and identification of EG-VEGF/prokineticins as cognate ligands for two orphan G-protein-coupled receptors. Biochem. Biophys. Res. Commun. 293, 396–402 - PubMed
-
- Lin D. C., Bullock C. M., Ehlert F. J., Chen J. L., Tian H., Zhou Q. Y. (2002) Identification and molecular characterization of two closely related G-protein-coupled receptors activated by prokineticins/endocrine gland vascular endothelial growth factor. J. Biol. Chem. 277, 19276–19280 - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
