

Excitatory GABA: How a Correct Observation May Turn Out to be an Experimental Artifact
- PMID: 22529813
- PMCID: PMC3329772
- DOI: 10.3389/fphar.2012.00065
Excitatory GABA: How a Correct Observation May Turn Out to be an Experimental Artifact
Abstract
The concept of the excitatory action of GABA during early development is based on data obtained mainly in brain slice recordings. However, in vivo measurements as well as observations made in intact hippocampal preparations indicate that GABA is in fact inhibitory in rodents at early neonatal stages. The apparent excitatory action of GABA seems to stem from cellular injury due to the slicing procedure, which leads to accumulation of intracellular Cl(-) in injured neurons. This procedural artifact was shown to be attenuated through various manipulations such as addition of energy substrates more relevant to the in vivo situation. These observations question the very concept of excitatory GABA in immature neuronal networks.
Keywords: GABA; brain slices; energy substrates; giant depolarizing potentials; in vivo versus in vitro.
Figures
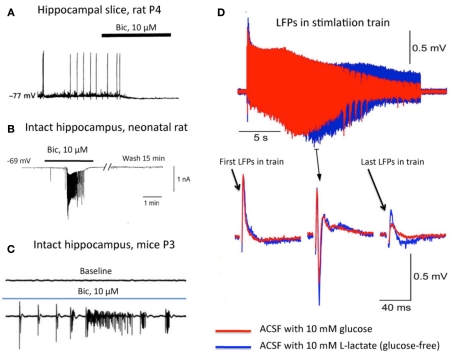
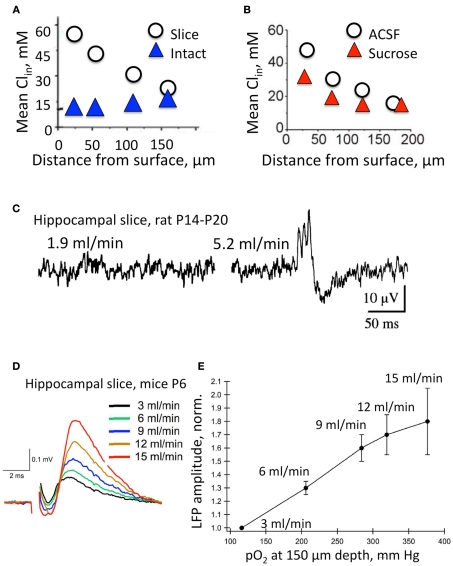
 ) and acute hippocampal slice preparations (◯) at P5–P7. Note the highly elevated Cl concentrations in neurons from the surface layers in the slice preparation (Modified from Dzhala et al., 2012). (B) The effects of slicing conditions on intracellular Cl concentration. Mean Cli as a function of depth in the hippocampal slices prepared from P5–P7 mice in control ACSF and in a high sucrose solution (Modified from Dzhala et al., 2012). (C–E) Genesis of network events and amplitude of local field potentials strongly depend upon the flow rate of ACSF. (C) Spontaneous network activity recorded at a low flow rate of 1.9 ml/min (left), and a high flow rate of 5.2 ml/min (right). Note sharp wave–ripple activity only at a high flow rate. Juvenile (P14–P20) transverse hippocampal 400–450 μm thick slices from Wistar rats were used here (from Hájos et al., 2009). (D) Examples of local field potentials measured in the same slice and electrode positions at different flow rates. Note the remarkable increase in amplitude when the flow rate is increased. (E) Summary of the dependence of local field potential (LFP) amplitudes on the oxygen levels and perfusion rates. Slices 400 μm thick from P4–P7 Swiss mice (from Ivanov et al., 2011).
) and acute hippocampal slice preparations (◯) at P5–P7. Note the highly elevated Cl concentrations in neurons from the surface layers in the slice preparation (Modified from Dzhala et al., 2012). (B) The effects of slicing conditions on intracellular Cl concentration. Mean Cli as a function of depth in the hippocampal slices prepared from P5–P7 mice in control ACSF and in a high sucrose solution (Modified from Dzhala et al., 2012). (C–E) Genesis of network events and amplitude of local field potentials strongly depend upon the flow rate of ACSF. (C) Spontaneous network activity recorded at a low flow rate of 1.9 ml/min (left), and a high flow rate of 5.2 ml/min (right). Note sharp wave–ripple activity only at a high flow rate. Juvenile (P14–P20) transverse hippocampal 400–450 μm thick slices from Wistar rats were used here (from Hájos et al., 2009). (D) Examples of local field potentials measured in the same slice and electrode positions at different flow rates. Note the remarkable increase in amplitude when the flow rate is increased. (E) Summary of the dependence of local field potential (LFP) amplitudes on the oxygen levels and perfusion rates. Slices 400 μm thick from P4–P7 Swiss mice (from Ivanov et al., 2011).References
-
- Abi-Saab W. M., Maggs D. G., Jones T., Jacob R., Srihari V., Thompson J., Kerr D., Leone P., Krystal J. H., Spencer D. D., During M. J., Sherwin R. S. (2002). Striking differences in glucose and lactate levels between brain extracellular fluid and plasma in conscious human subjects: effects of hyperglycemia and hypoglycemia. J. Cereb. Blood Flow. Metab. 22, 271–279 - PubMed
LinkOut - more resources
Full Text Sources