

Disassembling iron availability to phytoplankton
- PMID: 22529839
- PMCID: PMC3328120
- DOI: 10.3389/fmicb.2012.00123
Disassembling iron availability to phytoplankton
Abstract
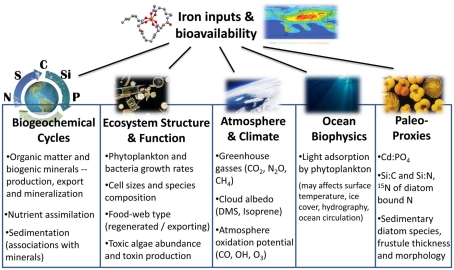
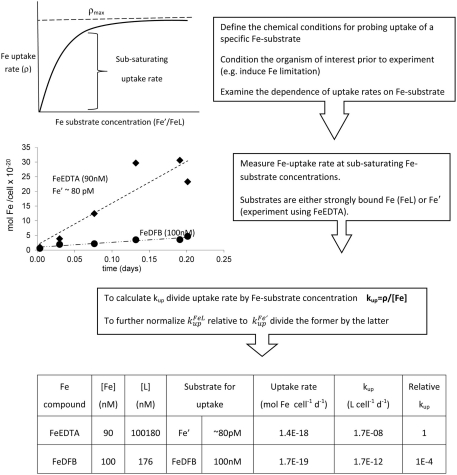
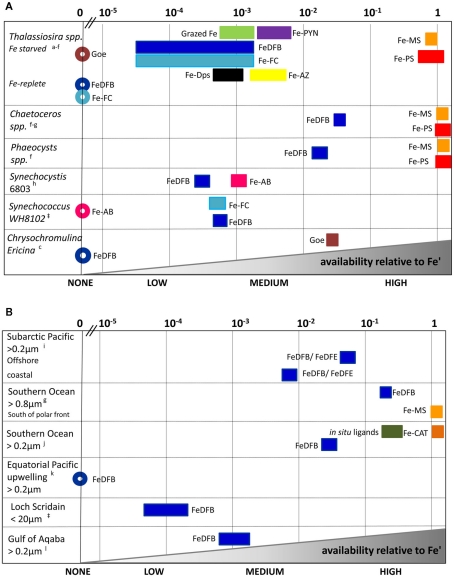
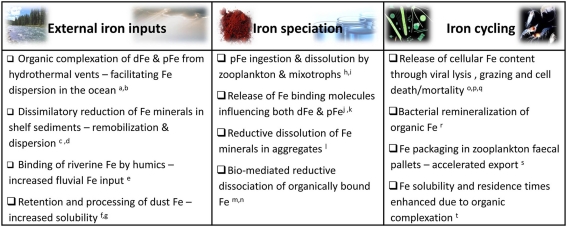
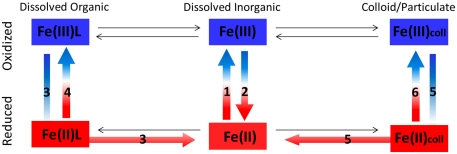
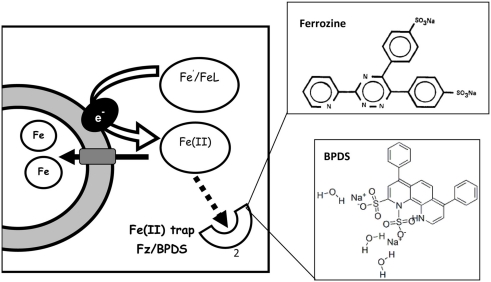
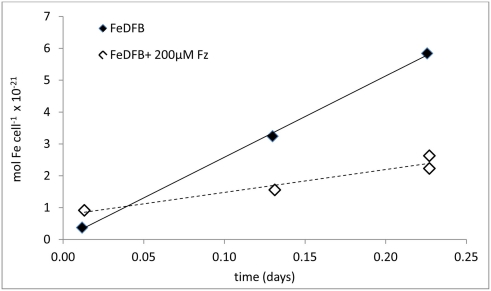
The bioavailability of iron to microorganisms and its underlying mechanisms have far reaching repercussions to many natural systems and diverse fields of research, including ocean biogeochemistry, carbon cycling and climate, harmful algal blooms, soil and plant research, bioremediation, pathogenesis, and medicine. Within the framework of ocean sciences, short supply and restricted bioavailability of Fe to phytoplankton is thought to limit primary production and curtail atmospheric CO(2) drawdown in vast ocean regions. Yet a clear-cut definition of bioavailability remains elusive, with elements of iron speciation and kinetics, phytoplankton physiology, light, temperature, and microbial interactions, to name a few, all intricately intertwined into this concept. Here, in a synthesis of published and new data, we attempt to disassemble the complex concept of iron bioavailability to phytoplankton by individually exploring some of its facets. We distinguish between the fundamentals of bioavailability - the acquisition of Fe-substrate by phytoplankton - and added levels of complexity involving interactions among organisms, iron, and ecosystem processes. We first examine how phytoplankton acquire free and organically bound iron, drawing attention to the pervasiveness of the reductive uptake pathway in both prokaryotic and eukaryotic autotrophs. Turning to acquisition rates, we propose to view the availability of various Fe-substrates to phytoplankton as a spectrum rather than an absolute "all or nothing." We then demonstrate the use of uptake rate constants to make comparisons across different studies, organisms, Fe-compounds, and environments, and for gaging the contribution of various Fe-substrates to phytoplankton growth in situ. Last, we describe the influence of aquatic microorganisms on iron chemistry and fate by way of organic complexation and bio-mediated redox transformations and examine the bioavailability of these bio-modified Fe species.
Keywords: bioavailability; biogeochemistry; iron; organic complexation; phytoplankton; redox reactions; speciation; uptake.
Figures


References
-
- Allen M. D., Del Campo J. A., Kropat J., Merchant S. S. (2007). FEA1, FEA2, and FRE1, encoding two homologous secreted proteins and a candidate ferrireductase, are expressed coordinately with FOX1 and FTR1 in iron-deficient Chlamydomonas reinhardtii. Eukaryotic Cell 6, 1841–185210.1128/EC.00205-07 - DOI - PMC - PubMed
-
- Chen M., Wang W. (2008). Accelerated uptake by phytoplankton of iron bound to humic acids. Aquat. Biol. 3, 155–16610.3354/ab00064 - DOI
-
- Eckhardt U., Buckhout T. J. (1998). Iron assimilation in Chlamydomonas reinhardtii involves ferric reduction and is similar to strategy I higher plants. J. Exp. Bot. 49, 1219–122610.1093/jexbot/49.324.1219 - DOI
LinkOut - more resources
Full Text Sources