

The differential production of cytokines by human Langerhans cells and dermal CD14(+) DCs controls CTL priming
- PMID: 22535664
- PMCID: PMC3382933
- DOI: 10.1182/blood-2011-08-371245
The differential production of cytokines by human Langerhans cells and dermal CD14(+) DCs controls CTL priming
Abstract
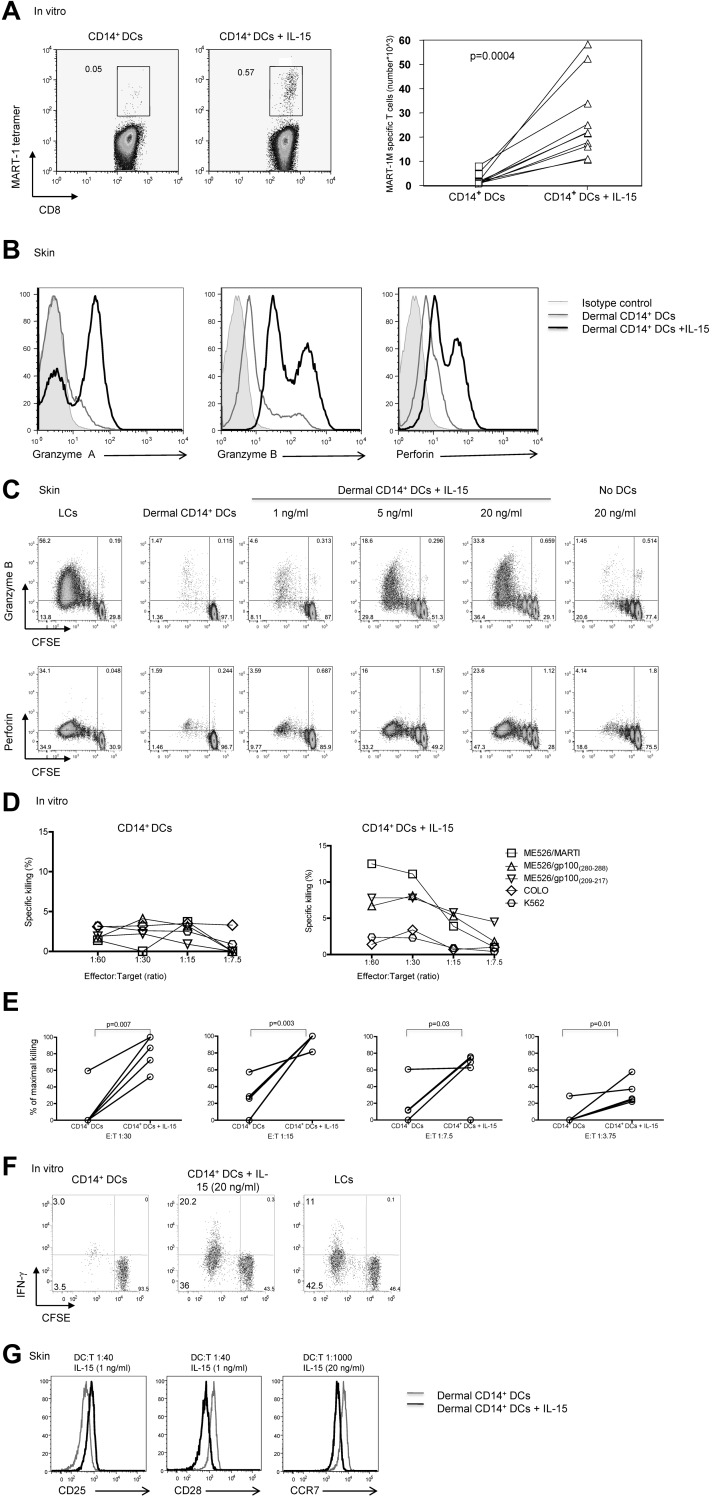
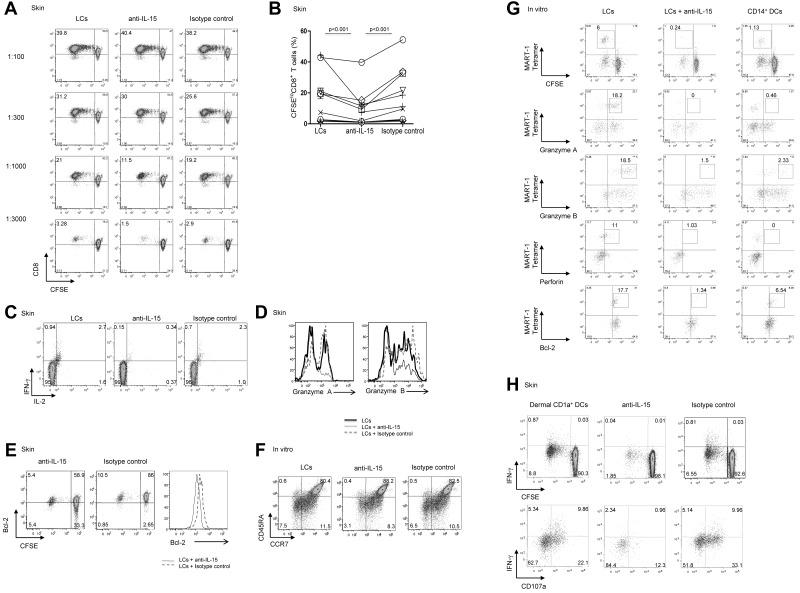
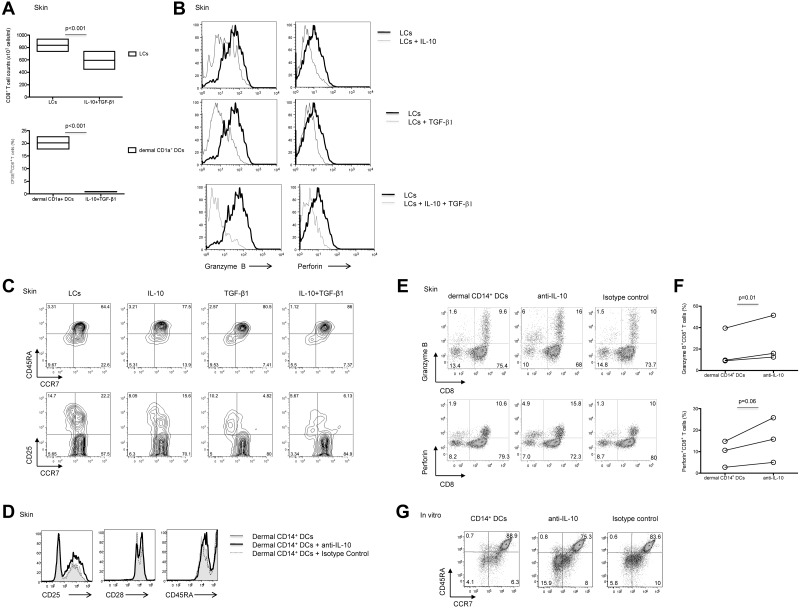
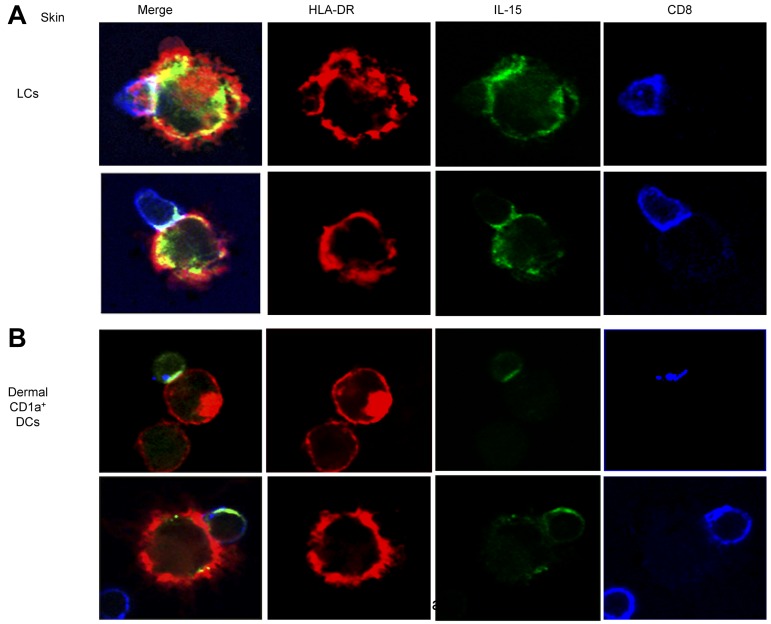
We recently reported that human epidermal Langerhans cells (LCs) are more efficient than dermal CD14(+) DCs at priming naive CD8(+) T cells into potent CTLs. We hypothesized that distinctive dendritic cell (DC) cytokine expression profiles (ie, IL-15 produced by LCs and IL-10 expressed by dermal CD14(+) DCs) might explain the observed functional difference. Blocking IL-15 during CD8(+) T-cell priming reduced T-cell proliferation by ∼ 50%. These IL-15-deprived CD8(+) T cells did not acquire the phenotype of effector memory cells. They secreted less IL-2 and IFN-γ and expressed only low amounts of CD107a, granzymes and perforin, and reduced levels of the antiapoptotic protein Bcl-2. Confocal microscopy analysis showed that IL-15 is localized at the immunologic synapse of LCs and naive CD8(+) T cells. Conversely, blocking IL-10 during cocultures of dermal CD14(+) DCs and naive CD8(+) T cells enhanced the generation of effector CTLs, whereas addition of IL-10 to cultures of LCs and naive CD8(+) T cells inhibited their induction. TGF-β1 that is transcribed by dermal CD14(+) DCs further enhanced the inhibitory effect of IL-10. Thus, the respective production of IL-15 and IL-10 explains the contrasting effects of LCs and dermal CD14(+) DCs on CD8(+) T-cell priming.
Figures
References
-
- Blattman JN, Greenberg PD. Cancer immunotherapy: a treatment for the masses. Science. 2004;305(5681):200–205. - PubMed
-
- Seder RA, Darrah PA, Roederer M. T-cell quality in memory and protection: implications for vaccine design. Nat Rev Immunol. 2008;8(4):247–258. - PubMed
-
- Pulendran B, Ahmed R. Translating innate immunity into immunological memory: implications for vaccine development. Cell. 2006;124(4):849–863. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
